Teleostei
| Teleostei Rentang waktu:
| |
|---|---|

| |
| Teleostei dari berbagai ordo, dilukis oleh Castelnau, 1856 (kiri ke kanan, atas ke bawah): Fistularia tabacaria (Syngnathiformes), Mylossoma duriventre (Characiformes), Mesonauta acora (Cichliformes), Corydoras splendens dan Pseudacanthicus spinosus (Siluriformes), Acanthurus coeruleus (Acanthuriformes), Stegastes pictus (Blenniiformes) | |
| Klasifikasi ilmiah | |
| Kerajaan: | Animalia |
| Filum: | Chordata |
| Kelas: | Actinopterygii |
| Infrakelas: | Teleosteomorpha |
| Divisi: | Teleostei J. P. Müller, 1845[3] |
| Subgrup | |
|
Lihat teks | |
Teleostei (bahasa Yunani teleios 'lengkap' + osteon 'tulang'), yang anggotanya dikenal sebagai teleosta,[4] sejauh ini adalah kelompok terbesar dari ikan bersirip kipas (kelas Actinopterygii),[a] mencakup 96% dari seluruh spesies ikan yang masih ada. Teleostei, yang secara bervariasi dianggap sebagai divisi atau infrakelas dalam sistem taksonomi yang berbeda, mencakup lebih dari 26.000 spesies yang disusun dalam sekitar 40 ordo dan 448 famili. Teleosta berkisar dari oarfish raksasa yang berukuran 76 m (249 ft) atau lebih, dan ikan matahari yang memiliki berat lebih dari 2 t (2,0 ton panjang; 2,2 ton pendek), hingga pejantan anglerfish Photocorynus spiniceps yang sangat kecil, hanya sepanjang 62 mm (2,44 in). Tidak hanya mencakup ikan berbentuk torpedo yang beradaptasi untuk kecepatan, tubuh teleosta dapat berbentuk pipih vertikal atau horizontal, silinder memanjang, atau memiliki bentuk khusus seperti pada anglerfish dan kuda laut.
Perbedaan antara teleosta dan ikan bertulang sejati lainnya terutama terletak pada tulang rahangnya; teleosta memiliki premaksila yang dapat digerakkan dan modifikasi terkait pada otot rahang yang memungkinkan mereka untuk menonjolkan rahangnya ke luar dari mulut. Hal ini merupakan keuntungan besar, yang memungkinkan mereka untuk menyambar mangsa dan menariknya ke dalam mulut. Pada teleosta yang lebih maju, premaksila yang membesar menjadi tulang utama penyangga gigi, dan maksila, yang melekat pada rahang bawah, bertindak sebagai tuas, mendorong dan menarik premaksila saat mulut dibuka dan ditutup. Tulang-tulang lain yang berada lebih jauh di belakang mulut berfungsi untuk menggiling dan menelan makanan. Perbedaan lainnya adalah lobus atas dan bawah dari sirip ekor (kaudal) memiliki ukuran yang kira-kira sama. Tulang belakang berakhir pada pangkal ekor, membedakan kelompok ini dari ikan lain di mana tulang belakang memanjang hingga ke lobus atas sirip ekor.
Teleosta telah mengadopsi berbagai strategi reproduksi. Sebagian besar menggunakan pembuahan eksternal: betina melepaskan sekumpulan telur, jantan membuahinya, dan larva berkembang tanpa keterlibatan induk lebih lanjut. Sejumlah besar teleosta adalah hermafrodit berurutan, memulai kehidupan sebagai betina dan beralih menjadi jantan pada tahap tertentu, dengan beberapa spesies yang membalikkan proses ini. Persentase kecil teleosta bersifat vivipar dan beberapa memberikan pengasuhan induk, biasanya ikan jantan yang menjaga sarang dan mengipasi telur untuk menjaga agar tetap mendapatkan oksigen yang cukup.
Teleosta memiliki nilai ekonomi penting bagi manusia, sebagaimana ditunjukkan oleh penggambaran mereka dalam seni selama berabad-abad. Industri perikanan memanen mereka untuk makanan, dan pemancing mencoba menangkap mereka sebagai olahraga. Beberapa spesies dibudidayakan secara komersial, dan metode produksi ini kemungkinan akan menjadi semakin penting di masa depan. Sementara jenis ikan Teleosta ainnya dipelihara di akuarium atau digunakan dalam penelitian, terutama di bidang genetika dan biologi perkembangan.
Anatomi

Ciri pembeda dari teleosta adalah premaksila yang dapat bergerak, lengkung saraf (arcus neuralis) yang memanjang di ujung sirip ekor, dan lempeng gigi basibranchial yang tidak berpasangan.[6] Premaksila tidak menempel pada neurokranium (kotak otak); tulang ini berperan dalam menjulurkan mulut dan menciptakan bukaan melingkar. Hal ini menurunkan tekanan di dalam mulut, sehingga mangsa terisap ke dalam. Rahang bawah dan maksila kemudian ditarik kembali untuk menutup mulut, dan ikan tersebut mampu mencengkeram mangsa. Sebaliknya, penutupan rahang biasa akan berisiko mendorong makanan keluar dari mulut. Pada teleosta yang lebih maju, premaksila membesar dan memiliki gigi, sedangkan maksila tidak bergigi. Maksila berfungsi untuk mendorong premaksila dan rahang bawah ke depan. Untuk membuka mulut, otot aduktor menarik bagian atas maksila ke belakang, mendorong rahang bawah ke depan. Selain itu, maksila sedikit berotasi, yang mendorong prosesus tulang ke depan yang saling mengunci dengan premaksila.[5]

Rahang faring teleosta, set rahang kedua yang terdapat di dalam tenggorokan, terdiri dari lima lengkung insang, yaitu lengkungan tulang yang menyangga insang. Tiga lengkung pertama mencakup satu basibranchial tunggal yang dikelilingi oleh dua hypobranchial, ceratobranchial, epibranchial, dan pharyngobranchial. Basibranchial median ditutupi oleh lempeng gigi. Lengkung keempat terdiri dari pasangan ceratobranchial dan epibranchial, dan terkadang tambahan beberapa pharyngobranchial dan sebuah basibranchial. Pangkal rahang faring bawah dibentuk oleh ceratobranchial kelima, sedangkan pharyngobranchial kedua, ketiga, dan keempat membentuk pangkal rahang atas. Pada teleosta yang lebih basal, rahang faring terdiri dari bagian-bagian tipis yang terpisah dengan jelas yang menempel pada neurokranium, gelang bahu, dan batang hioid. Fungsinya terbatas hanya untuk mengangkut makanan, dan mereka sebagian besar mengandalkan aktivitas rahang faring bawah. Pada teleosta yang lebih maju, rahangnya lebih kuat, dengan ceratobranchial kiri dan kanan menyatu menjadi satu rahang bawah; pharyngobranchial menyatu membentuk rahang atas besar yang bersendi dengan neurokranium. Mereka juga telah mengembangkan otot yang memungkinkan rahang faring berperan dalam menggiling makanan selain mengangkutnya.[7]
Sirip ekor bersifat homoserkal, yang berarti lobus atas dan bawah memiliki ukuran yang kira-kira sama. Tulang belakang berakhir pada pangkal ekor (pedunkula kaudal), yaitu dasar sirip ekor, yang membedakan kelompok ini dari kelompok di mana tulang belakang memanjang hingga ke lobus atas sirip ekor, seperti kebanyakan ikan dari zaman Paleozoikum (541 hingga 252 juta tahun yang lalu). Lengkung saraf memanjang membentuk uroneural yang memberikan penyangga bagi lobus atas ini.[5]
Teleosta cenderung lebih cepat dan lebih fleksibel dibandingkan ikan bertulang sejati yang lebih basal. Struktur rangka mereka telah berevolusi menjadi lebih ringan. Meskipun tulang teleosta terkalsifikasi dengan baik, tulang tersebut terbentuk dari kerangka penopang, bukan tulang spongiosa padat seperti pada ikan holostei. Selain itu, rahang bawah teleosta tereduksi menjadi hanya tiga tulang; dentari, tulang angular dan tulang artikular.[8] Saluran kelamin dan saluran kemih berakhir di belakang anus pada papila genital; hal ini diamati untuk menentukan jenis kelamin teleosta.[9]
Evolusi dan filogeni
Hubungan eksternal
Teleosta pertama kali diakui sebagai kelompok yang terpisah oleh ahli iktiologi Jerman Johannes Peter Müller pada tahun 1845.[10] Namanya berasal dari bahasa Yunani Kuno teleios, "lengkap" + osteon, "tulang".[11] Müller mendasarkan klasifikasi ini pada karakteristik jaringan lunak tertentu, yang kemudian terbukti bermasalah, karena tidak memperhitungkan ciri-ciri pembeda dari fosil teleosta. Pada tahun 1966, Greenwood dkk. memberikan klasifikasi yang lebih kuat.[10][12] Fosil tertua dari teleosteomorph (kelompok batang tempat teleosta kemudian berevolusi) berasal dari periode Trias (Prohalecites, Pholidophorus).[13][14] Namun, telah dikemukakan bahwa teleosta mungkin pertama kali berevolusi pada era Paleozoikum.[15] Selama era Mesozoikum dan Kenozoikum, mereka mengalami diversifikasi yang luas, dan hasilnya, 96% dari semua spesies ikan yang hidup saat ini adalah teleosta.[16]
Kladogram di bawah ini menunjukkan hubungan evolusi teleosta dengan klad ikan bertulang sejati lainnya yang masih ada,[15] dan dengan vertebrata berkaki empat (tetrapoda) yang berevolusi dari kelompok ikan bertulang sejati yang berkerabat selama periode Devon.[17][18]Perkiraan waktu divergensi (dalam jutaan tahun, jtl) berasal dari Near dkk., 2012.[15]
| Euteleostomi/ |
| ||||||||||||||||||||||||
| Osteichthyes |
Hubungan internal
Filogeni teleosta telah lama menjadi subjek perdebatan panjang, tanpa adanya konsensus mengenai filogeni mereka maupun waktu kemunculan kelompok-kelompok utamanya sebelum penerapan analisis kladistik berbasis DNA modern. Near dkk. (2012) meneliti filogeni dan waktu divergensi setiap garis keturunan utama, dengan menganalisis urutan DNA dari 9 gen yang tidak saling terkait pada 232 spesies. Mereka memperoleh filogeni yang terpecahkan dengan baik dengan dukungan yang kuat pada nodusnya (sehingga, pola percabangan yang ditampilkan kemungkinan besar benar). Mereka mengkalibrasi (menetapkan nilai aktual untuk) waktu percabangan dalam pohon ini dari 36 pengukuran waktu absolut yang dapat diandalkan dari catatan fosil.[15] Teleosta dibagi menjadi klad-klad utama yang ditunjukkan pada kladogram berikut,[19] beserta tanggalnya, mengikuti Near dkk.[15]Penelitian yang lebih baru membagi teleosta menjadi dua kelompok besar: Eloposteoglossocephala (Elopomorpha + Osteoglossomorpha) dan Clupeocephala (sisa teleosta lainnya).[20][21]
| Teleostei |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 310 jtl |

Kelompok ikan teleosta yang paling beragam saat ini adalah Percomorpha, yang antara lain mencakup tuna, kuda laut, gobi, cichlid, ikan pipih, wrasse, perch, anglerfish, dan ikan buntal.[22] Teleosta, dan khususnya percomorpha, berkembang pesat selama era Kenozoikum. Bukti fosil menunjukkan bahwa terjadi peningkatan besar dalam ukuran dan kelimpahan teleosta segera setelah peristiwa kepunahan massal pada batas Kapur-Paleogen sekitar 66 jtl.[23]

Tren evolusi

Fosil pertama yang dapat dikaitkan dengan kelompok yang beragam ini muncul pada Trias Awal,[24] setelah itu teleosta mengakumulasi bentuk tubuh baru terutama secara bertahap selama 150 juta tahun pertama evolusi mereka (Trias Awal hingga Kapur Awal).[24]
Teleosta yang masih hidup paling basal adalah Elopomorpha (sidat dan kerabatnya) dan Osteoglossomorpha (ikan gajah dan kerabatnya). Terdapat 800 spesies elopomorpha. Mereka memiliki larva berbentuk daun tipis yang dikenal sebagai leptocephali, yang terspesialisasi untuk lingkungan laut. Di antara elopomorpha, sidat memiliki tubuh memanjang dengan gelang panggul dan rusuk yang hilang serta elemen yang menyatu di rahang atas. Sekitar 200 spesies osteoglossomorpha didefinisikan oleh elemen tulang di lidah. Elemen ini memiliki basibranchial di belakangnya, dan kedua struktur tersebut memiliki gigi besar yang berpasangan dengan gigi pada parasfenoid di langit-langit mulut. Klad Otocephala mencakup Clupeiformes (haring) dan Ostariophysi (karper, lele, dan kerabatnya). Clupeiformes terdiri dari 350 spesies haring dan ikan mirip haring yang masih hidup. Kelompok ini dicirikan oleh sisik perut yang tidak biasa dan susunan hypural yang berbeda. Pada sebagian besar spesies, gelembung renang memanjang hingga ke kotak otak dan berperan dalam pendengaran. Ostariophysi, yang mencakup sebagian besar ikan air tawar, meliputi spesies yang telah mengembangkan beberapa adaptasi unik.[5] Salah satunya adalah Aparatus Weber, susunan tulang (osikel Weber) yang menghubungkan gelembung renang ke telinga bagian dalam. Hal ini meningkatkan pendengaran mereka, karena gelombang suara membuat gelembung bergetar, dan tulang-tulang tersebut mentransmisikan getaran ke telinga bagian dalam. Mereka juga memiliki sistem alarm kimia; ketika seekor ikan terluka, zat peringatan akan masuk ke dalam air, memperingatkan ikan-ikan di dekatnya.[25]
Mayoritas spesies teleosta termasuk dalam klad Euteleostei, yang terdiri dari 17.419 spesies yang diklasifikasikan dalam 2.935 genus dan 346 famili. Ciri-ciri bersama euteleostei meliputi kesamaan dalam perkembangan embrio struktur tulang atau tulang rawan yang terletak di antara kepala dan sirip punggung (tulang supraneural), pertumbuhan berlebih pada tulang stegural (tulang yang terletak di dekat lengkung saraf ekor), dan tulang rawan median kaudal yang terletak di antara hypural pada pangkal ekor. Mayoritas euteleostei berada dalam klad Neoteleostei. Ciri turunan dari neoteleostei adalah otot yang mengendalikan rahang faring, yang memberi mereka peran dalam menggiling makanan. Di dalam neoteleostei, anggota Acanthopterygii memiliki sirip punggung berduri yang berada di depan sirip punggung jari-jari lunak.[26] Sirip ini membantu memberikan daya dorong dalam lokomosi[27] dan mungkin juga berperan dalam pertahanan. Acanthomorpha telah mengembangkan sisik ctenoid berduri (berlawanan dengan sisik cycloid pada kelompok lain), premaksila bergigi, dan adaptasi yang lebih besar untuk berenang dengan kecepatan tinggi.[5]
Sirip adiposa, yang terdapat pada lebih dari 6.000 spesies teleosta, sering dianggap berevolusi sekali dalam garis keturunan dan telah hilang berkali-kali karena fungsinya yang terbatas. Sebuah studi tahun 2014 menantang gagasan ini dan menyarankan bahwa sirip adiposa adalah contoh evolusi konvergen. Pada Characiformes, sirip adiposa berkembang dari pertumbuhan berlebih setelah reduksi lipatan sirip larva, sedangkan pada Salmoniformes, sirip tersebut tampaknya merupakan sisa dari lipatan tersebut.[28]
Keanekaragaman

Terdapat lebih dari 26.000 spesies teleosta, dalam sekitar 40 ordo dan 448 famili,[29] menyusun 96% dari seluruh spesies ikan yang masih ada.[16] Sekitar 12.000 dari total 26.000 spesies ditemukan di habitat air tawar.[30] Teleosta ditemukan di hampir setiap lingkungan akuatik dan telah mengembangkan spesialisasi untuk makan dengan berbagai cara sebagai karnivora, herbivora, pemakan saring, dan parasit.[31] Teleosta terpanjang adalah oarfish raksasa, yang dilaporkan berukuran 76 m (249 ft) dan lebih,[32] tetapi ukuran ini masih kalah jauh dibandingkan dengan Leedsichthys yang telah punah, di mana satu individu diperkirakan memiliki panjang 276 m (906 ft).[33] Teleosta terberat diyakini sebagai ikan matahari, dengan spesimen yang didaratkan pada tahun 2003 memiliki perkiraan berat 23 t (23 ton panjang; 25 ton pendek),[34] sementara dewasa matang yang terkecil adalah pejantan anglerfish Photocorynus spiniceps yang hanya berukuran 62 mm (2,44 in), meskipun betinanya yang berukuran 50 mm (2 in) jauh lebih besar.[32] Dan Paedocypris progenetica adalah salah satu vertebrata terkecil.[35]

Ikan perairan terbuka biasanya berbentuk ramping seperti torpedo untuk meminimalkan turbulensi saat mereka bergerak di dalam air. Ikan karang hidup dalam lanskap bawah air yang kompleks dan relatif terbatas, dan bagi mereka, manuverabilitas lebih penting daripada kecepatan, dan banyak dari mereka telah mengembangkan tubuh yang mengoptimalkan kemampuan untuk melesat dan mengubah arah. Banyak yang memiliki tubuh terkompresi secara lateral (pipih dari sisi ke sisi) yang memungkinkan mereka masuk ke dalam celah dan berenang melalui celah sempit; beberapa menggunakan sirip dada mereka untuk lokomosi dan yang lain menggelombangkan sirip punggung dan dubur mereka.[36] Beberapa ikan telah menumbuhkan apendiks dermal (kulit) untuk kamuflase; leatjer-jacket berduri hampir tidak terlihat di antara rumput laut yang menyerupainya dan scorpionfish berumbai bersembunyi tak terlihat di dasar laut siap untuk menyergap mangsa. Beberapa seperti butterflyfish mata empat memiliki bintik mata untuk mengejutkan atau menipu, sementara yang lain seperti lepu memiliki pewarnaan aposematik untuk memperingatkan bahwa mereka beracun atau memiliki duri berbisa.[37]
Ikan pipih adalah ikan demersal (ikan pemakan dasar) yang menunjukkan tingkat asimetri yang lebih besar daripada vertebrata lainnya. Larvanya pada awalnya simetris bilateral tetapi mereka mengalami metamorfosis selama perkembangan mereka, dengan satu mata bermigrasi ke sisi lain kepala, dan mereka secara bersamaan mulai berenang dengan sisi tubuh mereka. Ini memiliki keuntungan bahwa, ketika mereka berbaring di dasar laut, kedua mata berada di atas, memberi mereka bidang pandang yang luas. Sisi atas biasanya berbintik-bintik dan bercak untuk kamuflase, sedangkan sisi bawahnya pucat.[38]
Beberapa teleosta adalah parasit. Remora memiliki sirip punggung depan yang termodifikasi menjadi pengisap besar yang mereka gunakan untuk menempel pada hewan inang seperti paus, penyu, hiu, atau pari, tetapi ini mungkin merupakan pengaturan komensal daripada parasit karena remora dan inang mendapat manfaat dari penghapusan ektoparasit dan serpihan kulit yang lepas.[39] Yang lebih berbahaya adalah lele parasit yang masuk ke ruang insang ikan dan memakan darah serta jaringannya.[40] Belut hidung pesek, meskipun biasanya merupakan pemakan bangkai, kadang-kadang melubangi daging ikan, dan telah ditemukan di dalam jantung seekor hiu mako sirip pendek.[41]
Beberapa spesies, seperti sidat listrik, dapat menghasilkan arus listrik yang kuat, cukup kuat untuk memingsankan mangsa.Ikan lain, seperti ikan pisau, menghasilkan dan merasakan medan listrik lemah untuk mendeteksi mangsa mereka; mereka berenang dengan punggung lurus untuk menghindari distorsi medan listrik mereka. Arus ini dihasilkan oleh otot atau sel saraf yang termodifikasi.[25]
-
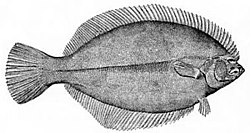 Flounder musim dingin bersifat asimetris, dengan kedua mata terletak di sisi kepala yang sama.
Flounder musim dingin bersifat asimetris, dengan kedua mata terletak di sisi kepala yang sama. -

-
 Ikan pisau Gymnarchus niloticus menghasilkan medan listrik lemah yang memungkinkannya untuk mendeteksi dan menemukan mangsa di air keruh.
Ikan pisau Gymnarchus niloticus menghasilkan medan listrik lemah yang memungkinkannya untuk mendeteksi dan menemukan mangsa di air keruh.



Persebaran
Teleosta tersebar di seluruh dunia dan menghuni sebagian besar lingkungan akuatik, termasuk lautan hangat dan dingin, air tawar yang mengalir maupun tenang, dan bahkan, dalam kasus pupfish gurun, di badan air yang terisolasi dan terkadang panas serta asin di gurun.[42][43] Keanekaragaman teleosta menjadi rendah di garis lintang yang sangat tinggi; di Franz Josef Land, hingga 82°LU, tutupan es serta suhu air di bawah 0 °C (32 °F) sepanjang sebagian besar tahun membatasi jumlah spesies; 75 persen spesies yang ditemukan di sana merupakan endemik Arktik.[44]

Dari kelompok utama teleosta, Elopomorpha, Clupeomorpha, dan Percomorpha (perch, tuna, dan banyak lainnya) semuanya memiliki persebaran di seluruh dunia dan terutama hidup di laut; Ostariophysi dan Osteoglossomorpha tersebar di seluruh dunia tetapi terutama hidup di air tawar, di mana kelompok yang terakhir disebutkan terutama berada di daerah tropis; Atherinomorpha (guppy, dll.) memiliki persebaran di seluruh dunia, baik di air tawar maupun asin, namun merupakan penghuni permukaan. Sebaliknya, Esociformes (pike) terbatas pada air tawar di Belahan Bumi Utara, sementara Salmoniformes (salmon, trout) ditemukan di zona iklim sedang baik di Utara maupun Selatan di air tawar, dengan beberapa spesies yang melakukan migrasi dari dan ke laut. Paracanthopterygii (kod, dll.) adalah ikan Belahan Bumi Utara, dengan spesies yang hidup di air asin maupun air tawar.[43]
Beberapa teleosta bersifat migratori; spesies air tawar tertentu berpindah di dalam sistem sungai setiap tahun; spesies lain bersifat anadromous, menghabiskan hidup mereka di laut dan bergerak ke pedalaman untuk memijah, dengan salmon dan bass bergaris sebagai contohnya. Lainnya, yang dicontohkan oleh sidat, bersifat katadromous, melakukan hal sebaliknya.[45] Sidat Eropa air tawar bermigrasi melintasi Samudra Atlantik saat dewasa untuk berkembang biak di antara rumput laut yang mengapung di Laut Sargasso. Ikan dewasa memijah di sini dan kemudian mati, tetapi anak-anaknya yang sedang berkembang terbawa oleh Arus Teluk menuju Eropa. Pada saat mereka tiba, mereka adalah ikan kecil yang memasuki estuaria dan menaiki sungai, mengatasi rintangan di jalur mereka untuk mencapai sungai kecil dan kolam tempat mereka menghabiskan kehidupan dewasa mereka.[46]
Teleosta termasuk trout cokelat dan osman bersisik ditemukan di danau pegunungan di Kashmir pada ketinggian hingga 3.819 m (12.530 ft).[47] Teleosta ditemukan pada kedalaman ekstrem di samudra; hadal snailfish telah terlihat pada kedalaman 7.700 m (25.300 ft), dan spesies terkait (yang belum diberi nama) telah terlihat pada kedalaman 8.145 m (26.720 ft).[48][49]
Fisiologi
Respirasi

Mekanisme respirasi utama pada teleosta, seperti pada kebanyakan ikan lainnya, adalah pertukaran gas melalui permukaan insang saat air ditarik masuk melalui mulut dan dipompa keluar melewati insang.
Selain gelembung renang, yang berisi sejumlah kecil udara, tubuh tidak memiliki cadangan oksigen, dan respirasi harus berlangsung terus-menerus sepanjang hidup ikan. Beberapa teleosta mengeksploitasi habitat di mana ketersediaan oksigen rendah, seperti air yang tergenang atau lumpur basah; mereka telah mengembangkan jaringan dan organ aksesori untuk mendukung pertukaran gas di habitat ini.[50]
Beberapa genus teleosta secara independen telah mengembangkan kemampuan bernapas di udara, dan beberapa telah menjadi amfibi. Beberapa blenni gigi sisir muncul ke permukaan untuk mencari makan di darat, dan sidat air tawar mampu menyerap oksigen melalui kulit yang lembap. Tembakul dapat bertahan di luar air untuk waktu yang cukup lama, menukarkan gas melalui kulit dan membran mukosa di mulut dan faring. Belut sawah memiliki lapisan mulut dengan vaskularisasi yang sama baiknya, dan dapat bertahan di luar air selama berhari-hari serta masuk ke keadaan istirahat (estivasi) di dalam lumpur.[51] Kelompok anabantoid telah mengembangkan struktur pernapasan aksesori yang dikenal sebagai organ labirin pada lengkung insang pertama dan ini digunakan untuk respirasi di udara
[Image of labyrinth organ in fish] , dan lele bernapas udara memiliki organ suprabranchial yang serupa. Ikan lele tertentu lainnya, seperti Loricariidae, mampu bernapas melalui udara yang ditahan di dalam saluran pencernaan mereka.[52]
Sistem sensorik

Teleosta memiliki organ sensorik yang sangat berkembang. Hampir semua ikan siang hari memiliki penglihatan warna setidaknya sebaik manusia normal. Banyak ikan juga memiliki kemoreseptor yang bertanggung jawab atas indra perasa dan penciuman yang tajam. Sebagian besar ikan memiliki reseptor sensitif yang membentuk sistem gurat sisi, yang mendeteksi arus dan getaran lembut, serta merasakan gerakan ikan dan mangsa di dekatnya.[53] Ikan merasakan suara dengan berbagai cara, menggunakan gurat sisi, gelembung renang, dan pada beberapa spesies menggunakan aparatus Weber. Ikan mengarahkan diri mereka menggunakan penanda lokasi, dan mungkin menggunakan peta mental berdasarkan berbagai penanda atau simbol. Eksperimen dengan labirin menunjukkan bahwa ikan memiliki memori spasial yang diperlukan untuk membuat peta mental semacam itu.[54]
Osmoregulasi

Kulit teleosta sebagian besar tidak permeabel terhadap air, dan antarmuka utama antara tubuh ikan dan lingkungannya adalah insang. Di air tawar, ikan teleosta menyerap air melalui insang mereka secara osmosis, sementara di air laut mereka kehilangan air. Demikian pula, garam berdifusi keluar melalui insang di air tawar dan masuk ke dalam di air asin. Flounder Eropa menghabiskan sebagian besar hidupnya di laut tetapi sering bermigrasi ke estuaria dan sungai. Di laut dalam satu jam, ia dapat memperoleh ion Na+ setara dengan empat puluh persen dari total kandungan natrium bebasnya, dengan 75 persen di antaranya masuk melalui insang dan sisanya melalui minum. Sebaliknya, di sungai hanya terjadi pertukaran dua persen dari kandungan Na+ tubuh per jam. Selain mampu secara selektif membatasi pertukaran garam dan air melalui difusi, terdapat mekanisme aktif di seluruh insang untuk eliminasi garam di air laut dan penyerapannya di air tawar.[55]
Termoregulasi
Ikan bersifat berdarah dingin, dan secara umum suhu tubuhnya sama dengan suhu lingkungan sekitarnya. Mereka memperoleh dan kehilangan panas melalui kulit, serta mengatur sirkulasi mereka sebagai respons terhadap perubahan suhu air dengan meningkatkan atau mengurangi aliran darah ke insang. Panas metabolik yang dihasilkan di otot atau usus dengan cepat dilepaskan melalui insang, dengan darah dialihkan menjauhi insang selama paparan terhadap suhu dingin.[56] Karena ketidakmampuan relatif mereka untuk mengontrol suhu darah, sebagian besar teleosta hanya dapat bertahan hidup dalam rentang suhu air yang kecil.[57]
Spesies teleosta yang menghuni perairan yang lebih dingin memiliki proporsi asam lemak tak jenuh yang lebih tinggi dalam membran sel otak dibandingkan dengan ikan dari perairan yang lebih hangat, yang memungkinkan mereka mempertahankan fluiditas membran yang tepat di lingkungan tempat mereka hidup.[58] Ketika teraklimatisasi dengan suhu dingin, ikan teleosta menunjukkan perubahan fisiologis pada otot rangka yang mencakup peningkatan kepadatan mitokondria dan kapiler.[59] Hal ini mengurangi jarak difusi dan membantu produksi ATP aerobik, yang membantu mengkompensasi penurunan laju metabolisme yang terkait dengan suhu yang lebih dingin.
Tuna dan ikan samudra lain yang berenang cepat mempertahankan otot mereka pada suhu yang lebih tinggi daripada lingkungan mereka untuk lokomosi yang efisien.[60] Tuna mencapai suhu otot 11 °C (19 °F) atau bahkan lebih tinggi di atas lingkungan sekitarnya dengan memiliki sistem aliran balik di mana panas metabolik yang dihasilkan oleh otot dan berada dalam darah vena, menghangatkan darah arteri sebelum mencapai otot. Adaptasi lain dari tuna untuk kecepatan mencakup tubuh berbentuk spindel yang ramping, sirip yang dirancang untuk mengurangi gaya hambat,[60] dan otot dengan kandungan mioglobin yang tinggi, yang memberikan warna kemerahan dan membuat penggunaan oksigen menjadi lebih efisien.[61] Di wilayah kutub dan di laut dalam, di mana suhu berada beberapa derajat di atas titik beku, beberapa ikan besar, seperti ikan pedang, marlin, dan tuna, memiliki mekanisme pemanas yang meningkatkan suhu otak dan mata, memungkinkan mereka memiliki penglihatan yang jauh lebih baik daripada mangsa mereka yang berdarah dingin.[62]
Daya Apung

Tubuh teleosta lebih padat daripada air, sehingga ikan harus mengkompensasi perbedaan tersebut, atau mereka akan tenggelam. Ciri khas dari Actinopteri (Chondrostei, Holostei, dan teleosta) adalah adanya gelembung renang.[63][64] Awalnya hadir pada leluhur bersama terakhir dari teleosta, organ ini telah hilang secara independen setidaknya 30–32 kali dalam setidaknya 79 dari 425 famili teleosta di mana gelembung renang tidak ada pada satu atau lebih spesies. Ketidakhadiran ini sering terjadi pada ikan perenang cepat seperti tuna dan makarel.[65] Gelembung renang membantu ikan menyesuaikan daya apung mereka melalui manipulasi gas, yang memungkinkan mereka untuk tetap berada pada kedalaman air saat ini, atau naik dan turun tanpa harus membuang energi untuk berenang. Pada kelompok yang lebih primitif seperti beberapa minnow, gelembung renang terbuka (fisostoma) ke esofagus. Pada ikan di mana gelembung renang tertutup (fisoklista), kandungan gas dikontrol melalui rete mirabile, sebuah jaringan pembuluh darah yang berfungsi sebagai penukar gas arus balik antara gelembung renang dan darah. [66]
Lokomosi

Ikan teleosta yang khas memiliki tubuh ramping untuk berenang cepat, dan lokomosi umumnya disediakan oleh undulasi (gerakan bergelombang) lateral dari bagian paling belakang batang tubuh dan ekor, yang mendorong ikan tersebut menembus air. [67] Ada banyak pengecualian untuk metode lokomosi ini, terutama di mana kecepatan bukan tujuan utama; di antara bebatuan dan di terumbu karang, berenang lambat dengan kemampuan manuver yang hebat mungkin merupakan atribut yang diinginkan.[68] Sidat bergerak dengan menggoyangkan seluruh tubuh mereka. Hidup di antara lamun dan alga, kuda laut mengambil postur tegak dan bergerak dengan mengepakkan sirip dadanya, dan pipefish yang berkerabat dekat bergerak dengan menggetarkan sirip punggungnya yang memanjang. Gobi "melompat" di sepanjang substrat, menopang diri mereka sendiri dan mendorong diri dengan sirip dada mereka.[69] Tembakul bergerak dengan cara yang hampir sama di daratan.[70] Pada beberapa spesies, pengisap panggul memungkinkan mereka untuk memanjat, dan gobi air tawar Hawaii memanjat air terjun saat bermigrasi.[69] Gurnard memiliki tiga pasang jari-jari bebas pada sirip dada mereka yang memiliki fungsi sensorik tetapi juga dapat mereka gunakan untuk berjalan di sepanjang substrat.[71] Ikan terbang meluncurkan diri mereka ke udara dan dapat meluncur dengan sirip dada mereka yang membesar hingga ratusan meter.[72]
Produksi suara
Kemampuan memproduksi suara untuk komunikasi tampaknya telah berevolusi secara independen di beberapa garis keturunan teleosta.[73] Suara dihasilkan baik dengan stridulasi atau dengan menggetarkan gelembung renang. Pada Sciaenidae, otot yang menempel pada gelembung renang menyebabkannya berosilasi dengan cepat, menciptakan suara seperti tabuhan gendang. Lele laut, kuda laut, dan ikan gerot-gerot melakukan stridulasi dengan menggesekkan bagian rangka, gigi, atau duri secara bersamaan. Pada ikan-ikan ini, gelembung renang dapat bertindak sebagai resonator. Suara stridulasi sebagian besar berkisar antara 1000–4000 Hz, meskipun suara yang dimodifikasi oleh gelembung renang memiliki frekuensi lebih rendah dari 1000 Hz.[74][75]
Reproduksi dan siklus hidup
Kebanyakan spesies teleosta bersifat ovipar, melakukan fertilisasi eksternal dengan telur dan sperma dilepaskan ke dalam air untuk pembuahan. Fertilisasi internal terjadi pada 500 hingga 600 spesies teleosta tetapi lebih umum bagi Chondrichthyes dan banyak tetrapoda. Hal ini melibatkan jantan yang menginseminasi betina dengan organ intromittent.[76] Kurang dari satu dari sejuta telur yang dibuahi secara eksternal bertahan hidup untuk berkembang menjadi ikan dewasa, namun terdapat peluang bertahan hidup yang jauh lebih baik di antara keturunan anggota sekitar selusin famili yang bersifat vivipar. Pada kelompok ini, telur dibuahi secara internal dan ditahan di dalam tubuh betina selama perkembangan. Beberapa spesies ini, seperti ikan akuarium vivipar dalam famili Poeciliidae, bersifat ovovivipar; setiap telur memiliki kantung kuning telur yang memberi nutrisi pada embrio yang sedang berkembang, dan ketika ini terserap habis, telur menetas dan larva dikeluarkan ke kolom air. Spesies lain, seperti splitfin dalam famili Goodeidae, sepenuhnya vivipar, dengan embrio yang berkembang diberi nutrisi dari suplai darah induk melalui struktur mirip plasenta yang berkembang di dalam rahim. Oofagi dipraktikkan oleh beberapa spesies, seperti Nomorhamphus ebrardtii; induk meletakkan telur yang tidak dibuahi yang dimakan oleh larva yang sedang berkembang di dalam uterus, dan kanibalisme intrauterin telah dilaporkan pada beberapa ikan julung-julung.[77]
Terdapat dua strategi reproduksi utama teleosta; semelparitas dan iteroparitas. Pada strategi pertama, individu berkembang biak sekali setelah mencapai kematangan dan kemudian mati. Hal ini karena perubahan fisiologis yang menyertai reproduksi pada akhirnya menyebabkan kematian.[78] Salmon dari genus Oncorhynchus terkenal dengan ciri ini; mereka menetas di air tawar dan kemudian bermigrasi ke laut hingga empat tahun sebelum melakukan perjalanan kembali ke tempat kelahirannya di mana mereka memijah dan mati. Semelparitas juga diketahui terjadi pada beberapa sidat dan smelt. Mayoritas spesies teleosta memiliki iteroparitas, di mana individu dewasa dapat berkembang biak beberapa kali selama hidupnya.[79]
Identitas dan penentuan jenis kelamin

88 persen spesies teleosta bersifat gonokoristik, memiliki individu yang tetap jantan atau betina sepanjang kehidupan dewasa mereka. Jenis kelamin individu dapat ditentukan secara genetik seperti pada burung dan mamalia, atau secara lingkungan seperti pada reptil. Pada beberapa teleosta, baik genetika maupun lingkungan berperan dalam menentukan jenis kelamin.[80] Untuk spesies yang jenis kelaminnya ditentukan oleh genetika, hal ini dapat terjadi dalam tiga bentuk. Dalam penentuan jenis kelamin monofaktorial, lokus tunggal menentukan pewarisan jenis kelamin. Baik sistem penentuan jenis kelamin XY maupun sistem penentuan jenis kelamin ZW ada pada spesies teleosta. Beberapa spesies, seperti platyfish selatan, memiliki kedua sistem tersebut dan seekor jantan dapat ditentukan oleh XY atau ZZ tergantung pada populasinya.[81]
Penentuan jenis kelamin multifaktorial terjadi pada banyak spesies Neotropis dan melibatkan sistem XY dan ZW. Sistem multifaktorial melibatkan penyusunan ulang kromosom seks dan autosom. Sebagai contoh, darter characine memiliki sistem multifaktorial ZW di mana betina ditentukan oleh ZW1W2 dan jantan oleh ZZ. Ikan serigala memiliki sistem multifaktorial XY di mana betina ditentukan oleh X1X1X2X2 dan jantan oleh X1X2Y.[82] Beberapa teleosta, seperti ikan zebra, memiliki sistem polifaktorial, di mana terdapat beberapa gen yang berperan dalam menentukan jenis kelamin.[83] Penentuan jenis kelamin yang bergantung pada lingkungan telah didokumentasikan pada setidaknya 70 spesies teleosta. Suhu adalah faktor utama, tetapi tingkat pH, laju pertumbuhan, kepadatan, dan lingkungan sosial juga dapat berperan. Bagi silverside Atlantik, pemijahan di perairan yang lebih dingin menghasilkan lebih banyak betina, sedangkan perairan yang lebih hangat menghasilkan lebih banyak jantan.[84]
Hermafroditisme
Beberapa spesies teleosta bersifat hermafrodit, yang dapat muncul dalam dua bentuk: simultan dan sekuensial. Pada bentuk pertama, spermatozoa dan telur keduanya ada di dalam gonad. Hermafroditisme simultan biasanya terjadi pada spesies yang hidup di kedalaman laut, di mana pasangan potensial tersebar jarang.[85][86] Pembuahan sendiri jarang terjadi dan hanya tercatat pada dua spesies, Kryptolebias marmoratus dan Kryptolebias hermaphroditus.[86] Dengan hermafroditisme sekuensial, individu dapat berfungsi sebagai satu jenis kelamin di awal kehidupan dewasanya dan berganti di kemudian hari. Spesies dengan kondisi ini termasuk ikan kakatua, wrasse, kerapu, ikan kepala rata, ikan bream laut dan lightfish.[85]
Protandri adalah ketika individu bermula sebagai jantan dan menjadi betina, sedangkan kondisi sebaliknya dikenal sebagai protogini, di mana yang terakhir lebih umum terjadi. Perubahan jenis kelamin dapat terjadi dalam berbagai konteks. Pada wrasse pembersih, di mana jantan memiliki harem hingga sepuluh betina, jika jantan diambil, betina terbesar dan paling dominan mengembangkan perilaku seperti jantan dan akhirnya memiliki testis. Jika dia diambil, betina peringkat berikutnya mengambil tempatnya. Pada spesies Anthias squamipinnis, di mana individu berkumpul dalam kelompok besar dan betina jauh melebihi jantan, jika sejumlah jantan diambil dari kelompok, jumlah betina yang sama akan berganti kelamin dan menggantikan mereka. Pada ikan badut, individu hidup dalam kelompok dan hanya dua yang terbesar dalam kelompok yang berkembang biak: betina terbesar dan jantan terbesar. Jika betina mati, jantan berganti jenis kelamin dan jantan terbesar berikutnya mengambil tempatnya.[87]
Pada anglerfish laut dalam (sub-ordo Ceratioidei), jantan yang jauh lebih kecil menempel secara permanen pada betina dan berdegenerasi menjadi lampiran penghasil sperma. Betina dan jantan yang menempel padanya menjadi "unit semi-hermafrodit".[88]
Taktik kawin

Terdapat beberapa sistem perkawinan yang berbeda di antara teleosta. Beberapa spesies bersifat promiskuitas, di mana jantan dan betina berkembang biak dengan banyak pasangan dan tidak ada pilihan pasangan yang jelas. Hal ini telah tercatat pada haring Baltik, guppy, kerapu Nassau, damselfish humbug, cichlid, dan wrasse kreol. Poligami, di mana satu jenis kelamin memiliki banyak pasangan, dapat muncul dalam berbagai bentuk. Poliandri terdiri dari satu betina dewasa yang berkembang biak dengan beberapa jantan, yang hanya berkembang biak dengan betina tersebut. Hal ini jarang terjadi di antara teleosta, dan ikan pada umumnya, tetapi ditemukan pada ikan badut. Selain itu, hal ini mungkin juga ada sampai batas tertentu di antara anglerfish, di mana beberapa betina memiliki lebih dari satu jantan yang menempel pada mereka. Poligini, di mana satu jantan berkembang biak dengan beberapa betina, jauh lebih umum. Hal ini tercatat pada sculpin, sunfish, darter, damselfish, dan cichlid di mana beberapa betina mungkin mengunjungi pejantan teritorial yang menjaga dan merawat telur serta anak-anaknya. Poligini juga dapat melibatkan seekor jantan yang menjaga harem yang terdiri dari beberapa betina. Hal ini terjadi pada spesies terumbu karang, seperti damselfish, wrasse, ikan kakatua, ikan dokter, ikan triger, dan tilefish.[79]
Pembiakan lek, di mana pejantan berkumpul untuk memamerkan diri kepada betina, telah tercatat setidaknya pada satu spesies, Cyrtocara eucinostomus. Sistem pembiakan menyerupai lek juga telah tercatat pada beberapa spesies lain. Pada spesies monogami, jantan dan betina dapat membentuk ikatan pasangan dan berkembang biak secara eksklusif dengan pasangannya. Hal ini terjadi pada lele air tawar Amerika Utara, banyak butterflyfish, kuda laut, dan beberapa spesies lainnya.[79] Percumbuan pada teleosta berperan dalam pengenalan spesies, memperkuat ikatan pasangan, posisi lokasi pemijahan, dan sinkronisasi pelepasan gamet. Ini termasuk perubahan warna, produksi suara, dan tampilan visual (ereksi sirip, berenang cepat, melompat ke permukaan), yang sering dilakukan oleh pejantan. Percumbuan dapat dilakukan oleh betina untuk mengatasi pejantan teritorial yang jika tidak dirayu akan mengusirnya.[89]

Dimorfisme seksual ada pada beberapa spesies. Individu dari satu jenis kelamin, biasanya jantan, mengembangkan karakteristik seksual sekunder yang meningkatkan peluang keberhasilan reproduksi mereka. Pada ikan lemadang, jantan memiliki kepala yang lebih besar dan tumpul dibandingkan betina. Pada beberapa spesies minnow, jantan mengembangkan kepala yang bengkak dan benjolan kecil yang dikenal sebagai tuberkel perkawinan selama musim kawin.[90] Pejantan kakatua bonggol hijau memiliki dahi yang lebih berkembang dengan "bubungan yang terosifikasi" yang berperan dalam adu kepala ritualistik.[91] Dimorfisme juga dapat berupa perbedaan pewarnaan. Sekali lagi, biasanya jantanlah yang berwarna cerah; pada killifish, ikan pelangi, dan wrasse, warnanya bersifat permanen, sedangkan pada spesies seperti minnow, stickleback, darter, dan sunfish, warnanya berubah seiring musim. Pewarnaan seperti itu bisa sangat mencolok bagi predator, menunjukkan bahwa dorongan untuk bereproduksi bisa lebih kuat daripada dorongan untuk menghindari predasi.[90]
Jantan yang tidak berhasil merayu betina mungkin mencoba mencapai keberhasilan reproduksi dengan cara lain. Pada spesies sunfish, seperti bluegill, jantan yang lebih besar dan lebih tua yang dikenal sebagai jantan parental, yang telah berhasil merayu betina, membangun sarang untuk telur yang mereka buahi. Jantan satelit yang lebih kecil meniru perilaku dan pewarnaan betina untuk mengakses sarang dan membuahi telur. Jantan lain, yang dikenal sebagai jantan penyusup (sneaker males), bersembunyi di dekatnya dan kemudian dengan cepat melesat ke sarang, membuahi sambil berlari. Jantan-jantan ini lebih kecil dari jantan satelit. Jantan penyusup juga ada pada salmon Oncorhynchus, di mana jantan kecil yang tidak mampu menempati posisi di dekat betina akan melesat masuk saat jantan dominan yang besar sedang memijah dengan betina.[92]
Lokasi pemijahan dan pengasuhan induk

Teleosta dapat memijah di kolom air atau, lebih umum lagi, di substrat. Pemijah kolom air sebagian besar terbatas pada terumbu karang; ikan akan bergegas menuju permukaan dan melepaskan gamet mereka. Hal ini tampaknya melindungi telur dari beberapa predator dan memungkinkan mereka menyebar luas melalui arus. Mereka tidak menerima pengasuhan induk. Pemijah kolom air lebih cenderung memijah dalam kelompok dibandingkan pemijah substrat. Pemijahan substrat umumnya terjadi di sarang, celah batu, atau bahkan liang. Beberapa telur dapat menempel pada berbagai permukaan seperti batu, tanaman, kayu, atau cangkang.[93]

Dari teleosta ovipar, sebagian besar (79 persen) tidak memberikan pengasuhan induk.[94] Pengasuhan oleh jantan jauh lebih umum daripada pengasuhan oleh betina.[94][95] Teritorialitas jantan "mempraadaptasi" suatu spesies untuk mengembangkan pengasuhan induk jantan.[96][97] Salah satu contoh pengasuhan induk betina yang tidak biasa terdapat pada ikan diskus, yang memberikan nutrisi bagi anak-anaknya yang sedang berkembang dalam bentuk lendir.[98] Beberapa spesies teleosta memiliki telur atau anak yang menempel pada atau dibawa di dalam tubuh mereka. Bagi lele laut, cardinalfish, jawfish, dan beberapa lainnya, telur dapat diinkubasi atau dibawa di dalam mulut, praktik yang dikenal sebagai eraman mulut. Pada beberapa cichlid Afrika, telur dapat dibuahi di sana. Pada spesies seperti acara bergaris, anak-anak dierami setelah menetas dan ini dapat dilakukan oleh kedua orang tua. Waktu pelepasan anak bervariasi antar spesies; beberapa pengeram mulut melepaskan anak yang baru menetas sementara yang lain mungkin menahannya sampai mereka menjadi juvenil. Selain eraman mulut, beberapa teleosta juga mengembangkan struktur untuk membawa anak. Pejantan nurseryfish memiliki kait tulang di dahi mereka untuk membawa telur yang telah dibuahi; telur-telur tersebut tetap berada di kait sampai menetas. Bagi kuda laut, pejantan memiliki kantong pengeraman tempat betina menyimpan telur yang telah dibuahi dan mereka tetap di sana sampai menjadi juvenil yang bebas berenang. Betina lele banjo memiliki struktur di perut mereka tempat telur menempel.[99]
Pada beberapa spesies yang mengasuh, anak-anak dari kelompok pemijahan sebelumnya mungkin tinggal bersama orang tua mereka dan membantu merawat anak-anak yang baru. Hal ini diketahui terjadi pada sekitar 19 spesies cichlid di Danau Tanganyika. Para pembantu ini mengambil bagian dalam membersihkan dan mengipasi telur serta larva, membersihkan lubang pembiakan, dan melindungi wilayah. Mereka mengalami penurunan laju pertumbuhan tetapi mendapatkan perlindungan dari predator. Parasitisme induk juga ada di antara teleosta; minnow dapat memijah di sarang sunfish serta sarang spesies minnow lainnya. Lele kuku dikenal meletakkan telur di substrat saat cichlid pengeram mulut mengumpulkan telur mereka, dan lele muda akan memakan larva cichlid tersebut. Kanibalisme filial terjadi pada beberapa famili teleosta dan mungkin berevolusi untuk melawan kelaparan.[100]
Pertumbuhan dan perkembangan
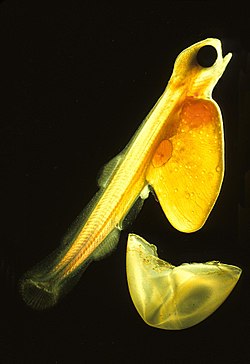
Teleosta memiliki empat tahapan kehidupan utama: telur, larva, juvenil, dan dewasa. Spesies dapat memulai kehidupan di lingkungan pelagis atau lingkungan demersal (dekat dasar laut). Sebagian besar teleosta laut memiliki telur pelagis, yang ringan, transparan, dan mengapung dengan selubung tipis. Telur pelagis mengandalkan arus laut untuk menyebar dan tidak menerima pengasuhan induk. Saat menetas, larvanya bersifat planktonik dan tidak mampu berenang. Mereka memiliki kantung kuning telur yang melekat pada tubuh yang menyediakan nutrisi. Sebagian besar spesies air tawar menghasilkan telur demersal yang tebal, berpigmen, relatif berat, dan mampu menempel pada substrat. Pengasuhan induk jauh lebih umum di antara ikan air tawar. Tidak seperti kerabat pelagis mereka, larva demersal mampu berenang dan makan segera setelah menetas.[85] Larva teleosta sering terlihat sangat berbeda dari bentuk dewasanya, terutama pada spesies laut. Beberapa larva bahkan sempat dianggap sebagai spesies yang berbeda dari bentuk dewasanya. Larva memiliki tingkat kematian yang tinggi; sebagian besar mati karena kelaparan atau predasi dalam minggu pertama mereka. Seiring pertumbuhan mereka, tingkat kelangsungan hidup meningkat, disertai dengan toleransi dan sensitivitas fisiologis, serta kompetensi ekologis dan perilaku yang lebih besar.[101]
Pada tahap juvenil, teleosta terlihat lebih mirip dengan bentuk dewasanya. Pada tahap ini, rangka aksial, organ dalam, sisik, pigmentasi, dan siripnya telah berkembang sepenuhnya. Transisi dari larva ke juvenil bisa singkat dan cukup sederhana, berlangsung dalam hitungan menit atau jam seperti pada beberapa damselfish, sementara pada spesies lain, seperti salmon, ikan tupai, gobi, dan ikan pipih, transisi tersebut lebih kompleks dan memakan waktu beberapa minggu untuk selesai.[102] Pada tahap dewasa, teleosta mampu menghasilkan gamet yang viabel untuk reproduksi. Seperti banyak ikan, teleosta terus tumbuh sepanjang hidup mereka. Umur panjang bergantung pada spesiesnya, dengan beberapa ikan buruan seperti perch Eropa dan bass mulut besar yang hidup hingga 25 tahun. Rockfish tampaknya menjadi teleosta yang paling panjang umur dengan beberapa spesies hidup lebih dari 100 tahun.[103]
Kawanan dan sekolah ikan

Banyak teleosta membentuk kawanan (shoal), yang memiliki berbagai tujuan pada spesies yang berbeda. Berenang dalam formasi (schooling) terkadang merupakan adaptasi antipredator, yang menawarkan kewaspadaan yang lebih baik terhadap predator. Sering kali lebih efisien untuk mengumpulkan makanan dengan bekerja sebagai kelompok, dan individu ikan mengoptimalkan strategi mereka dengan memilih untuk bergabung atau meninggalkan kawanan. Ketika predator diketahui, ikan mangsa merespons secara defensif, menghasilkan perilaku kawanan kolektif seperti gerakan yang tersinkronisasi. Respons tidak hanya terdiri dari upaya untuk bersembunyi atau melarikan diri; taktik antipredator mencakup misalnya memencar dan berkumpul kembali. Ikan juga berkumpul dalam kawanan untuk memijah.[104]
Hubungan dengan manusia
Kepentingan ekonomi

Teleosta memiliki kepentingan ekonomi dalam berbagai cara. Mereka ditangkap untuk makanan di seluruh dunia. Sejumlah kecil spesies seperti haring, kod, pollock, ikan teri, tuna, dan makarel menyediakan jutaan ton makanan bagi manusia setiap tahun, sementara banyak spesies lain ditangkap dalam jumlah yang lebih kecil.[105] Mereka menyumbang sebagian besar ikan yang ditangkap untuk olahraga.[106] Perikanan komersial dan rekreasi secara bersama-sama menyediakan lapangan kerja bagi jutaan orang.[107]
Sejumlah kecil spesies produktif termasuk karper, salmon,[108] nila, dan lele dibudidayakan secara komersial, menghasilkan jutaan ton makanan kaya protein setiap tahun. Pada tahun 2016, Organisasi Pangan dan Pertanian PBB memperkirakan produksi akan meningkat tajam sehingga pada tahun 2030, mungkin enam puluh dua persen ikan konsumsi akan berasal dari hasil budi daya.[109]
Ikan dikonsumsi dalam keadaan segar, atau dapat diawetkan dengan metode tradisional, yang meliputi kombinasi pengeringan, pengasapan, penggaraman, fermentasi,[110] dan pengacaran dalam cuka.[111] Metode pengawetan modern meliputi pembekuan, pengeringan beku, dan pemrosesan panas (seperti dalam proses pengalengan). Produk ikan beku meliputi filet yang dibalut tepung roti atau adonan, stik ikan, dan perkedel ikan. Tepung ikan digunakan sebagai suplemen makanan untuk ikan budi daya dan ternak. Minyak ikan dibuat baik dari hati ikan, yang sangat kaya akan Vitamin A dan Vitamin D, atau dari tubuh ikan berminyak seperti sarden dan haring, dan digunakan sebagai suplemen makanan serta untuk mengobati kekurangan vitamin.[112]
Beberapa spesies yang lebih kecil dan lebih berwarna berfungsi sebagai spesimen akuarium dan hewan peliharaan. Ikan serigala digunakan dalam industri kulit. Isinglass dibuat dari ikan kurau dan ikan gulamah.[106]
Dampak terhadap stok

Aktivitas manusia telah memengaruhi stok banyak spesies teleosta, melalui penangkapan ikan berlebih,[113] polusi, dan pemanasan global. Di antara banyak kejadian yang tercatat, penangkapan ikan berlebih menyebabkan runtuhnya populasi kod Atlantik secara total di lepas pantai Newfoundland pada tahun 1992, yang berujung pada penutupan perikanan tersebut oleh Kanada untuk waktu yang tidak ditentukan.[114] Polusi, terutama di sungai dan sepanjang pesisir, telah membahayakan teleosta karena limbah, pestisida, dan herbisida yang masuk ke dalam air. Banyak polutan, seperti logam berat, organoklorin, dan karbamat mengganggu reproduksi teleosta, sering kali dengan mengacaukan sistem endokrin mereka. Pada ikan roach, polusi sungai telah menyebabkan kondisi interseks, di mana gonad individu mengandung sel yang dapat membuat gamet jantan (seperti spermatogonia) dan sel yang dapat membuat gamet betina (seperti oogonia). Karena gangguan endokrin juga memengaruhi manusia, teleosta digunakan sebagai indikator keberadaan bahan kimia tersebut di dalam air. Polusi air menyebabkan kepunahan lokal populasi teleosta di banyak danau Eropa utara pada paruh kedua abad ke-20.[115]
Efek perubahan iklim terhadap teleosta bisa sangat kuat namun kompleks. Sebagai contoh, peningkatan curah hujan musim dingin (hujan dan salju) dapat membahayakan populasi ikan air tawar di Norwegia, sedangkan musim panas yang lebih hangat dapat meningkatkan pertumbuhan ikan dewasa.[116] Di lautan, teleosta mungkin mampu mengatasi pemanasan, karena hal itu hanyalah perpanjangan dari variasi alami iklim.[117] Masih belum pasti bagaimana pengasaman laut, yang disebabkan oleh kenaikan kadar karbon dioksida, akan memengaruhi teleosta.[118]
Interaksi lainnya

Beberapa teleosta berbahaya. Sebagian, seperti lele ekor sembilang (Plotosidae), ikan lepu (Scorpaenidae), atau ikan batu (Synanceiidae) memiliki duri berbisa yang dapat melukai serius atau membunuh manusia. Beberapa, seperti sidat listrik dan lele listrik, dapat memberikan sengatan listrik yang parah. Lainnya, seperti piranha dan barakuda, memiliki gigitan yang kuat dan terkadang menyerang manusia yang sedang mandi atau berenang.[106] Laporan menunjukkan bahwa beberapa famili lele dapat tumbuh cukup besar untuk memangsa manusia yang sedang mandi.
Medaka dan ikan zebra digunakan sebagai model penelitian untuk studi dalam genetika dan biologi perkembangan. Ikan zebra adalah vertebrata laboratorium yang paling umum digunakan,[106] menawarkan keuntungan berupa kesamaan genetik dengan mamalia, ukuran kecil, kebutuhan lingkungan yang sederhana, larva transparan yang memungkinkan pencitraan non-invasif, keturunan yang banyak, pertumbuhan cepat, dan kemampuan untuk menyerap mutagen yang ditambahkan ke dalam air mereka.[119]
Dalam seni
Ikan teleosta sering menjadi subjek dalam seni, mencerminkan kepentingan ekonomi mereka, setidaknya selama 14.000 tahun. Mereka umumnya dikerjakan dalam pola-pola di Mesir Kuno, memperoleh makna mitologis di Yunani Kuno dan Roma, dan dari sana masuk ke dalam Kekristenan sebagai simbol keagamaan; seniman di Tiongkok dan Jepang juga menggunakan citra ikan secara simbolis. Teleosta menjadi umum dalam Seni Renaisans, dengan lukisan benda mati mencapai puncak popularitas di Belanda pada abad ke-17. Pada abad ke-20, seniman yang berbeda seperti Klee, Magritte, Matisse, dan Picasso menggunakan representasi teleosta untuk mengekspresikan tema yang sangat berbeda, dari yang menarik hingga kekerasan.[120] Ahli zoologi dan seniman Ernst Haeckel melukis teleosta dan hewan lainnya dalam karyanya tahun 1904, Kunstformen der Natur. Haeckel telah diyakinkan oleh Goethe dan Alexander von Humboldt bahwa dengan membuat penggambaran akurat dari bentuk-bentuk alami yang asing, seperti dari laut dalam, ia tidak hanya dapat menemukan "hukum asal-usul dan evolusi mereka tetapi juga menekan ke bagian rahasia keindahan mereka dengan membuat sketsa dan melukis".[121]
-
 Lukisan dinding memancing, Makam Menna sang juru tulis, Thebes, Mesir Kuno, ca 1422–1411 SM
Lukisan dinding memancing, Makam Menna sang juru tulis, Thebes, Mesir Kuno, ca 1422–1411 SM -

-
 Lukisan Zaman Keemasan Belanda: Benda Mati Ikan dengan Lautan Badai, Willem Ormea dan Abraham Willaerts, 1636
Lukisan Zaman Keemasan Belanda: Benda Mati Ikan dengan Lautan Badai, Willem Ormea dan Abraham Willaerts, 1636 -
 Ikan Mandarin oleh Bian Shoumin, Dinasti Qing, abad ke-18
Ikan Mandarin oleh Bian Shoumin, Dinasti Qing, abad ke-18 -
 Saito Oniwakamaru melawan ikan mas raksasa di air terjun Bishimon oleh Utagawa Kuniyoshi, abad ke-19
Saito Oniwakamaru melawan ikan mas raksasa di air terjun Bishimon oleh Utagawa Kuniyoshi, abad ke-19 -
 Benda Mati dengan Makarel, Lemon dan Tomat, Vincent van Gogh, 1886
Benda Mati dengan Makarel, Lemon dan Tomat, Vincent van Gogh, 1886 -
 Ostraciontes oleh Ernst Haeckel, 1904. Sepuluh teleosta, dengan Lactoria cornuta di tengah.
Ostraciontes oleh Ernst Haeckel, 1904. Sepuluh teleosta, dengan Lactoria cornuta di tengah. -
 Sihir Ikan, Paul Klee, minyak dan cat air dipernis, 1925
Sihir Ikan, Paul Klee, minyak dan cat air dipernis, 1925








Catatan
- ^ Tiga kelompok lainnya adalah Holostei (bowfin dan gar), Chondrostei (sturgeon dan paddlefish), serta Cladistia (bichir dan reedfish).
Referensi
- ^ Palmer, Douglas (1999). The Marshall Illustrated Encyclopedia of Dinosaurs & Prehistoric Animals. Marshall Editions. ISBN 978-1-84028-152-1.
- ^ "The Paleobiology Database". The Paleobiology Database. 14 June 2013. Diarsipkan dari versi aslinya tanggal 27 March 2020. Diakses tanggal 14 June 2013.
- ^ Müller, Johannes (1845). "Über den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische". Archiv für Naturgeschichte. 11 (1): 129.
- ^ "teleost". Dictionary.com Unabridged. Random House.
- ^ a b c d e Benton, Michael (2005). "The Evolution of Fishes After the Devonian". Vertebrate Palaeontology (Edisi 3rd). John Wiley & Sons. hlm. 175–184. ISBN 978-1-4051-4449-0.
- ^ Patterson, C.; Rosen, D. E. (1977). "Review of ichthyodectiform and other Mesozoic teleost fishes, and the theory and practice of classifying fossils". Bulletin of the American Museum of Natural History. 158 (2): 81–172. hdl:2246/1224.
- ^ Vandewalle, P.; Parmentier, E.; Chardon, M. (2000). "The branchial basket in Teleost feeding" (PDF). Cybium. 24 (4): 319–342.
- ^ Bone, Q.; Moore, R. (2008). Biology of Fishes. Garland Science. hlm. 29. ISBN 978-0-415-37562-7.
- ^ Jamieson, Barrie G. M. (12 September 2019). Reproductive Biology and Phylogeny of Fishes, Vol 8B: Part B: Sperm Competition Hormones. CRC Press. ISBN 978-1-4398-4358-1.
- ^ a b Greenwood, P.; Rosen, D.; Weitzman, S.; Myers, G. (1966). "Phyletic studies of teleostean fishes, with a provisional classification of living forms". Bulletin of the American Museum of Natural History. 131: 339–456. hdl:2246/1678.
- ^ "Teleost". Merriam-Webster. Diakses tanggal 20 April 2016.
- ^ Arratia, G. (1998). "Basal teleosts and teleostean phylogeny: response to C. Patterson". Copeia. 1998 (4): 1109–1113. doi:10.2307/1447369. JSTOR 1447369.
- ^ Arratia, G. (2015). "Complexities of early teleostei and the evolution of particular morphological structures through time". Copeia. 103 (4): 999–1025. doi:10.1643/CG-14-184. S2CID 85808890.
- ^ Romano, Carlo; Koot, Martha B.; Kogan, Ilja; Brayard, Arnaud; Minikh, Alla V.; Brinkmann, Winand; Bucher, Hugo; Kriwet, Jürgen (February 2016). "Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution". Biological Reviews. 91 (1): 106–147. doi:10.1111/brv.12161. PMID 25431138. S2CID 5332637.
- ^ a b c d e Near, Thomas J.; et al. (2012). "Resolution of ray-finned fish phylogeny and timing of diversification". PNAS. 109 (34): 13698–13703. Bibcode:2012PNAS..10913698N. doi:10.1073/pnas.1206625109. PMC 3427055. PMID 22869754.
- ^ a b Berra, Tim M. (2008). Freshwater Fish Distribution. University of Chicago Press. hlm. 55. ISBN 978-0-226-04443-9.
- ^ Betancur-R., Ricardo; et al. (2013). "The Tree of Life and a New Classification of Bony Fishes". PLOS Currents: Tree of Life. 5 (Edisi 1st). doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288. hdl:2027.42/150563. PMC 3644299. PMID 23653398.
- ^ Laurin, M.; Reisz, R. R. (1995). "A reevaluation of early amniote phylogeny". Zoological Journal of the Linnean Society. 113 (2): 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x.
- ^ Betancur-R; et al. (2016). "Phylogenetic Classification of Bony Fishes Version 4". Deepfin. Diarsipkan dari asli tanggal 11 July 2017. Diakses tanggal 30 December 2016.
- ^ Study Resolves 50-Year Dispute of Teleost Fishes Ancestral Lineage
- ^ Genome structures resolve the early diversification of teleost fishes
- ^ Betancur-R, Ricardo; Wiley, Edward O.; Arratia, Gloria; Acero, Arturo; Bailly, Nicolas; Miya, Masaki; Lecointre, Guillaume; Ortí, Guillermo (6 July 2017). "Phylogenetic classification of bony fishes". BMC Evolutionary Biology. 17 (1): 162. Bibcode:2017BMCEE..17..162B. doi:10.1186/s12862-017-0958-3. PMC 5501477. PMID 28683774.
- ^ Sibert, E. C.; Norris, R. D. (2015-06-29). "New Age of Fishes initiated by the Cretaceous−Paleogene mass extinction". PNAS. 112 (28): 8537–8542. Bibcode:2015PNAS..112.8537S. doi:10.1073/pnas.1504985112. PMC 4507219. PMID 26124114.
- ^ a b Clarke, John T.; Friedman, Matt (August 2018). "Body-shape diversity in Triassic–Early Cretaceous neopterygian fishes: sustained holostean disparity and predominantly gradual increases in teleost phenotypic variety". Paleobiology. 44 (3): 402–433. Bibcode:2018Pbio...44..402C. doi:10.1017/pab.2018.8. S2CID 90207334.
- ^ a b Helfman, Collette, Facey and Bowen pp. 268–274
- ^ Helfman, Collette, Facey and Bowen pp. 274–276
- ^ Drucker, E. G.; Lauder, G. V. (2001). "Locomotor function of the dorsal fin in teleost fishes: experimental analysis of wake forces in sunfish". The Journal of Experimental Biology. 204 (Pt 17): 2943–2958. Bibcode:2001JExpB.204.2943D. doi:10.1242/jeb.204.17.2943. PMID 11551984.
- ^ Steward, T. A.; Smith, W. L.; Coates, M. I. (2014). "The origins of adipose fins: an analysis of homoplasy and the serial homology of vertebrate appendages". Proceedings of the Royal Society B. 281 (1781) 20133120. doi:10.1098/rspb.2013.3120. PMC 3953844. PMID 24598422.
- ^ Miller, Stephen; Harley, John P. (2007). Zoology (Edisi 7th). McGraw-Hill. hlm. 297.
- ^ Lackmann, Alec R.; Andrews, Allen H.; Butler, Malcolm G.; Bielak-Lackmann, Ewelina S.; Clark, Mark E. (2019-05-23). "Bigmouth Buffalo Ictiobus cyprinellus sets freshwater teleost record as improved age analysis reveals centenarian longevity". Communications Biology. 2 (1): 197. doi:10.1038/s42003-019-0452-0. PMC 6533251. PMID 31149641.
- ^ Dorit, R. L.; Walker, W. F.; Barnes, R. D. (1991). Zoology. Saunders College Publishing. hlm. 67–69. ISBN 978-0-03-030504-7.
- ^ a b Guinness World Records 2015. Guinness World Records. 2014. hlm. 60. ISBN 978-1-908843-70-8.
- ^ Martill, D.M. (1988). "Leedsichthys problematicus, a giant filter-feeding teleost from the Jurassic of England and France". Neues Jahrbuch für Geologie und Paläontologie. 1988 (11): 670–680. doi:10.1127/njgpm/1988/1988/670.
- ^ Roach, John (13 May 2003). "World's Heaviest Bony Fish Discovered?". National Geographic News. Diarsipkan dari asli tanggal 17 May 2003. Diakses tanggal 9 January 2016.
- ^ Kottelat, Maurice; Britz, Ralf; Hui, Tan Heok; Witte, Kai-Erik (2006-04-22). "Paedocypris, a new genus of Southeast Asian cyprinid fish with a remarkable sexual dimorphism, comprises the world's smallest vertebrate". Proceedings of the Royal Society B: Biological Sciences. 273 (1589): 895–899. doi:10.1098/rspb.2005.3419. PMC 1560243. PMID 16627273.
- ^ Maddock, L.; Bone, Q.; Rayner, J.M.V. (1994). The Mechanics and Physiology of Animal Swimming. Cambridge University Press. hlm. 54–56. ISBN 978-0-521-46078-1.
- ^ Ross, David A. (2000). The Fisherman's Ocean. Stackpole Books. hlm. 136–138. ISBN 978-0-8117-2771-6.
- ^ Schreiber, Alexander M. (2006). "Asymmetric craniofacial remodeling and lateralized behavior in larval flatfish". The Journal of Experimental Biology. 209 (Pt 4): 610–621. Bibcode:2006JExpB.209..610S. doi:10.1242/jeb.02056. PMID 16449556.
- ^ Jackson, John (30 November 2012). "How does the Remora develop its sucker?". National History Museum. Diakses tanggal 2 January 2016.
- ^ Combes, Claude (2001). Parasitism: The Ecology and Evolution of Intimate Interactions. University of Chicago Press. hlm. 23. ISBN 978-0-226-11446-0.
- ^ Caira, J.N.; Benz, G.W.; Borucinska, J.; Kohler, N.E. (1997). "Pugnose eels, Simenchelys parasiticus (Synaphobranchidae) from the heart of a shortfin mako, Isurus oxyrinchus (Lamnidae)". Environmental Biology of Fishes. 49 (1): 139–144. Bibcode:1997EnvBF..49..139C. doi:10.1023/a:1007398609346. S2CID 37865366.
- ^ Dudek and ICF International (2012). Desert Renewable Energy Conservation Plan (DRECP) Baseline Biology Report. California Energy Commission.
- ^ a b "Actinopterygii - ray-finned fishes". University College, London.
- ^ Chernova, N. V.; Friedlander, A. M.; Turchik, A.; Sala, E. (2014). "Franz Josef Land: extreme northern outpost for Arctic fishes". PeerJ. 2 e692. doi:10.7717/peerj.692. PMC 4266852. PMID 25538869.
- ^ "What is an anadromous fish? A catadromous fish?". Fish FAQ. NOAA. Diarsipkan dari asli tanggal 20 January 2016. Diakses tanggal 12 January 2016.
- ^ "Anguilla anguilla (Linnaeus, 1758)". Cultured Aquatic Species Information Programme. Organisasi Pangan dan Pertanian: Fisheries and Aquaculture Department. 1 January 2004. Diakses tanggal 2 January 2016.
- ^ Raina, H. S.; Petr, T. "Coldwater Fish and Fisheries in the Indian Himalayas: Lakes and Reservoirs". Food and Agriculture Organization. Diakses tanggal 6 January 2016.
- ^ Morelle, Rebecca (7 October 2008). "'Deepest ever' living fish filmed". BBC News. Diakses tanggal 5 February 2016.
- ^ Morelle, Rebecca (19 December 2014). "New record for deepest fish". BBC News. Diakses tanggal 5 February 2016.
- ^ Meurant, Gerard (1984). Fish Physiology V10A. Academic Press. hlm. 263–. ISBN 978-0-08-058531-4.
- ^ Liem, Karel F. (1998). Paxton, J.R.; Eschmeyer, W.N. (ed.). Encyclopedia of Fishes. Academic Press. hlm. 173–174. ISBN 978-0-12-547665-2.
- ^ Armbruster, Jonathan W. (1998). "Modifications of the digestive tract for holding air in loricariid and scoloplacid catfishes" (PDF). Copeia. 1998 (3): 663–675. doi:10.2307/1447796. JSTOR 1447796.
- ^ Orr, James (1999). Fish. Microsoft Encarta 99. ISBN 978-0-8114-2346-5.
- ^ Journal of Undergraduate Life Sciences. "Appropriate maze methodology to study learning in fish" (PDF). Diarsipkan dari asli (PDF) tanggal 6 July 2011. Diakses tanggal 28 May 2009.
- ^ Bentley, P.J. (2013). Endocrines and Osmoregulation: A Comparative Account in Vertebrates. Springer. hlm. 26. ISBN 978-3-662-05014-9.
- ^ Whittow, G. Causey (2013). Comparative Physiology of Thermoregulation: Special Aspects of Thermoregulation. Academic Press. hlm. 223. ISBN 978-1-4832-5743-3.
- ^ McFarlane, Paul (1 January 1999). "Warm-blooded fish". Monthly Bulletin. Hamilton and District Aquarium Society. Diarsipkan dari asli tanggal 15 May 2013. Diakses tanggal 6 January 2016.
- ^ Logue, J. A.; Vries, A. L. de; Fodor, E.; Cossins, A. R. (2000-07-15). "Lipid compositional correlates of temperature-adaptive interspecific differences in membrane physical structure". Journal of Experimental Biology. 203 (14): 2105–2115. Bibcode:2000JExpB.203.2105L. doi:10.1242/jeb.203.14.2105. PMID 10862723.
- ^ Johnston, I. A.; Dunn, J. (1987). "Temperature acclimation and metabolism in ectotherms with particular reference to teleost fish". Symposia of the Society for Experimental Biology. 41: 67–93. PMID 3332497.
- ^ a b Martin, R. Aidan (April 1992). "Fire in the Belly of the Beast". ReefQuest Centre for Shark Research. Diakses tanggal 6 January 2016.
- ^ Brown, W. Duane (1962). "The concentration of myoglobin and hemoglobin in tuna flesh". Journal of Food Science. 27 (1): 26–28. doi:10.1111/j.1365-2621.1962.tb00052.x.
- ^ Fritsches, Kerstin (11 January 2005). "Warm eyes give deep-sea predators super vision". University of Queensland. Diakses tanggal 6 January 2016.
- ^ Tatsumi, Norifumi; Kobayashi, Ritsuko; Yano, Tohru; Noda, Masatsugu; Fujimura, Koji; Okada, Norihiro; Okabe, Masataka (2016). "Molecular developmental mechanism in polypterid fish provides insight into the origin of vertebrate lungs". Scientific Reports. 6 30580. Bibcode:2016NatSR...630580T. doi:10.1038/srep30580. PMC 4964569. PMID 27466206.
- ^ Funk, E. C.; Breen, C.; Sanketi, B. D.; Kurpios, N.; McCune, A. (2020). "Changes in Nkx2.1, Sox2, Bmp4, and Bmp16 expression underlying the lung-to-gas bladder evolutionary transition in ray-finned fishes". Evolution & Development. 22 (5): 384–402. doi:10.1111/ede.12354. PMC 8013215. PMID 33463017.
- ^ McCune, Amy R.; Carlson, Rose L. (2004). "Twenty ways to lose your bladder: Common natural mutants in zebrafish and widespread convergence of swim bladder loss among teleost fishes". Evolution & Development. 6 (4): 246–259. doi:10.1111/j.1525-142X.2004.04030.x.
- ^ Kardong, K. (2008). Vertebrates: Comparative anatomy, function, evolution (Edisi 5th). Boston: McGraw-Hill. ISBN 978-0-07-304058-5.
- ^ Numerical Studies of Hydrodynamics of Fish Locomotion and Schooling by a Vortex Particle Method. 2008. hlm. 1–4. ISBN 978-1-109-14490-1.
- ^ Kapoor, B.G.; Khanna, Bhavna (2004). Ichthyology Handbook. Springer. hlm. 149–151. ISBN 978-3-540-42854-1.
- ^ a b Patzner, Robert; Van Tassell, James L.; Kovacic, Marcelo; Kapoor, B.G. (2011). The Biology of Gobies. CRC Press. hlm. 261, 507. ISBN 978-1-4398-6233-9.
- ^ Pace, C. M.; Gibb A. C. (2009). "Mudskipper pectoral fin kinematics in aquatic and terrestrial environments". The Journal of Experimental Biology. 212 (Pt 14): 2279–2286. Bibcode:2009JExpB.212.2279P. doi:10.1242/jeb.029041. PMID 19561218.
- ^ Jamon, M.; Renous, S.; Gasc, J.P.; Bels, V.; Davenport, J. (2007). "Evidence of force exchanges during the six-legged walking of the bottom-dwelling fish, Chelidonichthys lucerna". Journal of Experimental Zoology. 307 (9): 542–547. Bibcode:2007JEZA..307..542J. doi:10.1002/jez.401. PMID 17620306.
- ^ Dasilao, J.C.; Sasaki, K. (1998). "Phylogeny of the flyingfish family Exocoetidae (Teleostei, Beloniformes)". Ichthyological Research. 45 (4): 347–353. Bibcode:1998IchtR..45..347D. doi:10.1007/BF02725187. S2CID 24966029.
- ^ Rice, A. N.; et al. (2022). "Evolutionary Patterns in Sound Production across Fishes". Ichthyology & Herpetology. 110 (1): 1–12. doi:10.1643/i2020172. S2CID 245914602.
- ^ "How do fish produce sounds?". Discovery of Sound in the Sea. Diarsipkan dari asli tanggal 15 February 2017. Diakses tanggal 17 February 2017.
- ^ Lobel, P. S. "Fish Courtship and Mating Sounds". Massachusetts Institute of Technology. Diarsipkan dari asli tanggal 10 January 2018. Diakses tanggal 17 February 2017.
- ^ Wootton and Smith p. 5.
- ^ Springer, Joseph; Holley, Dennis (2012). An Introduction to Zoology. Jones & Bartlett Publishers. hlm. 370. ISBN 978-0-7637-5286-6.
- ^ Wootton and Smith p. 4.
- ^ a b c Helfman, Collette, Facey and Bowen p. 457
- ^ Wootton and Smith p. 2.
- ^ Wootton and Smith pp. 14, 19.
- ^ Wootton and Smith p. 20.
- ^ Wootton and Smith pp. 21–22.
- ^ Wootton and Smith p. 21–22.
- ^ a b c Laying, E. "Fish Reproduction" (PDF). Diarsipkan dari asli (PDF) tanggal 14 November 2014. Diakses tanggal 7 January 2016.
- ^ a b Wootton and Smith p. 2–4.
- ^ Helfman, Collette, Facey and Bowen p. 458
- ^ Wootton and Smith p. 320
- ^ Helfman, Collette, Facey and Bowen p. 465
- ^ a b Helfman, Collette, Facey and Bowen p. 463
- ^ Muñoz, R.; Zgliczynski, B.; Laughlin, J.; Teer, B. (2012). "Extraordinary aggressive behavior from the giant coral reef fish, Bolbometopon muricatum, in a remote marine reserve". PLOS ONE. 7 (6) e38120. Bibcode:2012PLoSO...738120M. doi:10.1371/journal.pone.0038120. PMC 3368943. PMID 22701606.
- ^ Helfman, Collette, Facey and Bowen p. 473
- ^ Helfman, Collette, Facey and Bowen p. 465–68
- ^ a b Reynolds, John; Nicholas B. Goodwin; Robert P. Freckleton (19 March 2002). "Evolutionary Transitions in Parental Care and Live Bearing in Vertebrates". Philosophical Transactions of the Royal Society B: Biological Sciences. 357 (1419): 269–281. doi:10.1098/rstb.2001.0930. PMC 1692951. PMID 11958696.
- ^ Clutton-Brock, T. H. (1991). The Evolution of Parental Care. Princeton, New Jersey: Princeton University Press.
- ^ Werren, John; Mart R. Gross; Richard Shine (1980). "Paternity and the evolution of male parentage". Journal of Theoretical Biology. 82 (4): 619–631. doi:10.1016/0022-5193(80)90182-4. PMID 7382520. Diakses tanggal 15 September 2013.
- ^ Baylis, Jeffrey (1981). "The Evolution of Parental Care in Fishes, with reference to Darwin's rule of male sexual selection". Environmental Biology of Fishes. 6 (2): 223–251. Bibcode:1981EnvBF...6..223B. doi:10.1007/BF00002788. S2CID 19242013.
- ^ Wootton and Smith p. 280
- ^ Wootton and Smith pp. 257–61
- ^ Helfman, Collette, Facey and Bowen pp. 472–73
- ^ Helfman, Collette, Facey and Bowen pp. 146–47
- ^ Helfman, Collette, Facey and Bowen pp. 149
- ^ Helfman, Collette, Facey and Bowen pp. 153–56
- ^ Pitcher, Tony J. (1986). "12. Functions of Shoaling Behaviour in Teleosts". The Behaviour of Teleost Fishes. Springer. hlm. 294–337. doi:10.1007/978-1-4684-8261-4_12. ISBN 978-1-4684-8263-8.
- ^ "Capture production by principal species in 2012" (PDF). Fishery and Aquaculture Statistics 2012. Food and Agriculture Organization. hlm. 12. Diakses tanggal 10 February 2016.
- ^ a b c d Kisia, S. M. (2010). Vertebrates: Structures and Functions. CRC Press. hlm. 22. ISBN 978-1-4398-4052-8.
- ^ "New Economic Report Finds Commercial and Recreational Saltwater Fishing Generated More Than Two Million Jobs". National Oceanic and Atmospheric Administration. Diakses tanggal 10 February 2016.
- ^ Scottish Fish Farm Production Survey 2014. The Scottish Government/Riaghaltas na h-Alba. September 2015. ISBN 978-1-78544-608-5.
- ^ "Fish to 2030: prospects for fisheries and aquaculture (Report 83177)". Food and Agriculture Organization; World Bank Group. 1 December 2013. hlm. 1–102. Diarsipkan dari asli tanggal 2 February 2016. Diakses tanggal 3 January 2016.
- ^ "Fish and fish products". Food and Agriculture Organization. Diarsipkan dari asli tanggal 8 February 2019. Diakses tanggal 8 April 2016.
- ^ Frank, Michele. "Pickled Fish". Diakses tanggal 6 July 2025.
- ^ Maqsood, Sajid; Singh, Prabjeet; Samoon, Munir Hassan; Wani, Gohar Bilal. "Various Fish and Fish Products Being Produced in Fish Processing Industries and Their Value Addition". Aquafind (Aquatic Fish Database). Diarsipkan dari asli tanggal 12 October 2011. Diakses tanggal 8 April 2016.
- ^ Vince, Gaia (20 September 2012). "How the world's oceans could be running out of fish". BBC. Diakses tanggal 1 May 2016.
- ^ Kunzig, R. (April 1995). "Twilight of the Cod". Discover: 52.
- ^ Wootton and Smith 2014, pp. 123–125
- ^ Kernan, Martin; Battarbee, Richard W.; Moss, Brian R. (2011). Climate Change Impacts on Freshwater Ecosystems. John Wiley & Sons. hlm. 93. ISBN 978-1-4443-9127-5.
- ^ Fisheries Management and Climate Change in the Northeast Atlantic Ocean and the Baltic Sea. Nordic Council of Ministers. 2008. hlm. 48. ISBN 978-92-893-1777-1.
- ^ Committee on the Review of the National Ocean Acidification Research and Monitoring Plan, Ocean Studies Board, Division on Earth and Life Studies, National Research Council (2013). Review of the Federal Ocean Acidification Research and Monitoring Plan. National Academies Press. hlm. 3. ISBN 978-0-309-30152-7. Pemeliharaan CS1: Banyak nama: authors list (link)
- ^ "Five reasons why zebrafish make excellent research models". NC3RS. 10 April 2014. Diakses tanggal 15 February 2016.
- ^ Moyle, Peter B.; Moyle, Marilyn A. (May 1991). "Introduction to fish imagery in art". Environmental Biology of Fishes. 31 (1): 5–23. Bibcode:1991EnvBF..31....5M. doi:10.1007/bf00002153. S2CID 33458630.
- ^ Richards, Robert J. "The Tragic Sense of Ernst Haeckel: His Scientific and Artistic Struggles" (PDF). University of Chicago. Diakses tanggal 30 April 2016.
Bibliografi
- Helfman, G.; Collette, B. B.; Facey, D. E.; Bowen, B. W. (2009). The Diversity of Fishes: Biology, Evolution, and Ecology (PDF) (Edisi 2nd). Wiley-Blackwell. ISBN 978-1-4051-2494-2. Diarsipkan dari asli (PDF) tanggal 26 April 2021. Diakses tanggal 18 January 2016.
- Wootton, Robert J.; Smith, Carl (2014). Reproductive Biology of Teleost Fishes. Wiley. ISBN 978-1-118-89139-1.
Pranala luar
 Media terkait Teleostei di Wikimedia Commons
Media terkait Teleostei di Wikimedia Commons Informasi terkait dengan Teleostei dari Wikispecies.
Informasi terkait dengan Teleostei dari Wikispecies.
Konten ini disalin dari wikipedia, mohon digunakan dengan bijak.

