Kosmoceratops
| Kosmoceratops Rentang waktu: Kapur Akhir,
| |
|---|---|

| |
| Cetakan tengkorak dengan rekonstruksi tulang predentari, tanduk kanan, dan bagian kiri wajah | |
| Klasifikasi ilmiah | |
| Kerajaan: | Animalia |
| Filum: | Chordata |
| Klad: | Dinosauria |
| Klad: | †Ornithischia |
| Klad: | †Neornithischia |
| Klad: | †Ceratopsia |
| Famili: | †Ceratopsidae |
| Subfamili: | †Chasmosaurinae |
| Genus: | †Kosmoceratops Sampson et al., 2010 |
| Spesies: | †K. richardsoni
|
| Nama binomial | |
| †Kosmoceratops richardsoni Sampson et al., 2010
| |
Kosmoceratops (/ˌkɒzməˈsɛrətɒps/[1]) adalah sebuah genus dinosaurus ceratopsid yang hidup di Amerika Utara sekitar 76–75,9 juta tahun yang lalu selama periode Kapur Akhir. Spesimen-spesimennya ditemukan di Utah dalam Formasi Kaiparowits di Monumen Nasional Grand Staircase–Escalante pada tahun 2006 dan 2007, termasuk sebuah tengkorak dewasa dan kerangka postkranial serta beberapa spesimen subdewasa parsial. Pada tahun 2010, spesimen dewasa tersebut dijadikan holotipe dari genus dan spesies baru Kosmoceratops richardsoni; nama generiknya berarti "wajah bertanduk berhias", dan nama spesifiknya menghormati Scott Richardson, yang menemukan spesimen-spesimen tersebut. Penemuan ini merupakan bagian dari gelombang penemuan ceratopsia pada awal abad ke-21, dan Kosmoceratops dianggap penting karena ornamentasi tengkoraknya yang rumit.
Kosmoceratops diperkirakan memiliki panjang 4,5 m (15 ft) dan berat 1,2 t (1,3 ton pendek). Sebagai seekor ceratopsid, ia merupakan hewan kuadrupedal dengan kerangka yang berkonstruksi kokoh. Hewan ini memiliki paruh berbentuk segitiga dengan ujung runcing dan tanduk hidung menyerupai bilah dengan bagian atas yang memipih. Sementara tanduk di atas matanya mengarah ke depan atau ke belakang pada sebagian besar kerabatnya, tanduk Kosmoceratops mengarah ke atas dan ke samping, kemudian ke bawah, serta berujung runcing. Jumbai lehernya berukuran pendek dari depan ke belakang, dengan fenestra parietal (bukaan yang menembus jumbai) kecil, dan sepuluh epiosifikasi menyerupai kail pada tepi belakangnya, dengan delapan melengkung ke depan dan dua melengkung ke samping. Dengan lima belas tanduk dan struktur menyerupai tanduk yang berkembang dengan baik, hewan ini memiliki tengkorak paling berhias dari semua spesies dinosaurus yang diketahui. Kosmoceratops adalah ceratopsid chasmosaurinae dan awalnya diusulkan berkerabat dekat dengan Vagaceratops (yang juga memiliki epiosifikasi yang melengkung ke depan pada bagian belakang jumbainya) namun hal ini telah diperdebatkan, dengan beberapa penulis mendapati bahwa Vagaceratops lebih dekat kekerabatannya dengan Chasmosaurus. Kosmoceratops juga dianggap berkerabat dekat dengan Spiclypeus, yang memiliki jumbai serupa.
Studi terhadap histologi tulang menunjukkan bahwa Kosmoceratops tumbuh dengan cepat dan memiliki tingkat metabolisme yang tinggi, mirip dengan burung dan mamalia modern. Gigi ceratopsid beradaptasi untuk memproses tanaman berserat; koprolit (kotoran fosil) dari Formasi Kaiparowits yang mengandung kayu mungkin dihasilkan oleh ceratopsid. Fungsi jumbai dan tanduk ceratopsia telah lama diperdebatkan, dan mencakup fungsi untuk peragaan visual, pertarungan, dan pengenalan spesies. Formasi Kaiparowits berasal dari akhir zaman Campanium dan diendapkan di Laramidia, sebuah benua pulau, ketika Amerika Utara terbelah di bagian tengah oleh Lajur Laut Interior Barat. Lingkungan ini didominasi oleh lahan basah dan mendukung fauna yang beragam, termasuk dinosaurus seperti chasmosaurinae Utahceratops. Didasarkan sebagian pada hubungan kekerabatan antara Kosmoceratops dan chasmosaurinae lain dari kurun waktu yang sama, telah diusulkan bahwa Laramidia terbagi menjadi "provinsi-provinsi" dinosaurus dengan spesies endemik terpisah (interpretasi ini menunjukkan bahwa Kosmoceratops di selatan berkerabat paling dekat dengan Vagaceratops di utara yang terpisah secara geografis), namun gagasan ini telah ditentang.
Penemuan

Sejak tahun 2000, Museum Sejarah Alam Utah (UMNH) dan Biro Pengelolaan Lahan telah melakukan survei paleontologi terhadap Formasi Kaiparowits di Monumen Nasional Grand Staircase–Escalante (GSENM) di Utah selatan. Monumen nasional ini didirikan pada tahun 1996 sebagian untuk pelestarian dan studi fosil-fosilnya, dan survei-survei di sana telah menghasilkan berbagai macam fosil dinosaurus yang unik. Kru lapangan dari berbagai institusi lain juga turut berpartisipasi, dan upaya kolaboratif ini dinamakan Proyek Cekungan Kaiparowits.[2][3] Di antara penemuan yang didapatkan terdapat tiga takson ceratopsia (dinosaurus bertanduk) baru, yang salah satunya diidentifikasi dari dua lokasi (Lokasi UMNH VP 890 dan 951) yang ditemukan oleh anggota kru lapangan sukarelawan Scott Richardson selama musim lapangan tahun 2006 dan 2007. Awalnya dinosaurus ini disebut sebagai "takson baru Kaiparowits A" dan diidentifikasi sebagai sebuah ceratopsid chasmosaurinae dalam sebuah buku terbitan tahun 2010.[3][4] Sebelum proyek ini, satu-satunya sisa ceratopsia yang ditemukan di formasi tersebut adalah gigi terisolasi yang tidak informatif.[5] Fosil-fosil yang digali diangkut ke UMNH, di mana bongkahan-bongkahan tersebut dipreparasi oleh para sukarelawan menggunakan jarum dan pahat udara pneumatik lalu disusun kembali.[2][6]
Pada tahun 2010, ahli paleontologi Scott D. Sampson beserta kolega menamai genus dan spesies baru Kosmoceratops richardsoni, dengan spesimen UMNH VP 17000 (dari Lokasi VP 890) sebagai holotipe. Nama generiknya berasal dari bahasa Yunani Kuno kosmos, yang berarti "berhias", dan ceratops, yang berarti "wajah bertanduk". Nama spesifiknya menghormati Richardson, yang menemukan holotipe tersebut beserta banyak fosil lainnya di GSENM.[2] Nama lengkapnya dapat diterjemahkan menjadi "wajah bertanduk berhias dari Richardson".[1] Holotipe ini mencakup sebuah tengkorak dewasa yang hampir utuh namun kehilangan tulang predentari (tulang paling depan dari rahang bawah) serta sebagian kecil wajah bagian kiri dan jumbai leher (bagian dari tulang jugal, skuamosal, dan parietal), dan moncongnya membengkok ke kanan akibat distorsi postmortem (setelah kematian). Sebagian besar kerangka aksial (seperti tulang belakang dan tulang rusuk) ditemukan berasosiasi dengan tengkorak tersebut, begitu juga dengan sebagian gelang panggul dan sebuah anggota gerak. Sebanyak 45 persen dari kerangka postkranial diperkirakan terawetkan, sebagian besarnya masih dalam tahap preparasi pada tahun 2010.[2][4][7] Spesimen yang ditetapkan sebagai spesies ini mencakup UMNH VP 16878,[3] sebuah tengkorak disartikulasi dari spesimen subdewasa (antara remaja dan dewasa) yang berukuran sekitar setengah dari individu dewasa, tanpa tulang premaksila, rostral, dan predentari, serta spesimen UMNH VP 21339, baik subdewasa maupun dewasa yang terdisartikulasi. Secara keseluruhan, dilaporkan ada empat spesimen yang ditemukan.[2][8][3]

Para pendeskripsi Kosmoceratops menamai genera chasmosaurinae baru Utahceratops (juga dari Formasi Kaiparowits) dan Vagaceratops (dari Formasi Dinosaur Park, yang spesies tunggalnya, C. irvinensis, sebelumnya ditempatkan ke dalam genus Chasmosaurus) pada artikel yang sama.[2] Genera ini, yang dianggap tidak biasa jika dibandingkan dengan anggota tipikal kelompoknya, merupakan bagian dari gelombang penemuan ceratopsia pada awal abad ke-21, saat banyak taksa baru dinamai (sebuah studi pada tahun 2013 menyatakan bahwa setengah dari seluruh genera yang valid dinamai sejak tahun 2003, dan dekade tersebut telah disebut sebagai "renaisans ceratopsid").[1][3][9] Fokus utama artikel yang ditulis oleh Sampson dan kolega adalah bagaimana ketiga genera ceratopsia ini memberikan bukti baru untuk merekonstruksi paleobiogeografi terkait ruang dan waktu mereka.[2] Dalam rilis pers tahun 2010 yang mengumumkan studi tersebut, Sampson mendeskripsikan Kosmoceratops sebagai "salah satu hewan paling menakjubkan yang pernah diketahui, dengan tengkorak raksasa yang dihiasi oleh bermacam-macam ornamen tulang", dan menganggap GSENM sebagai "salah satu kuburan dinosaurus besar terakhir di negara tersebut yang sebagian besar belum dieksplorasi".[1]

Pada tahun 2017, pemerintah AS mengumumkan rencana untuk menyusutkan ukuran monumen GSENM (menjadi sedikit lebih dari setengah ukurannya) dan Bears Ears guna memungkinkan penambangan batu bara dan pengembangan energi lainnya di lahan tersebut, yang mana ini merupakan pengurangan monumen nasional AS terbesar dalam sejarah.[10][11] Sampson, yang telah mengawasi sebagian besar penelitian awal di monumen sebelumnya, mengutarakan kekhawatirannya bahwa langkah tersebut akan mengancam penemuan-penemuan selanjutnya. Berbagai media massa menekankan pentingnya penemuan fosil di area tersebut—termasuk lebih dari 25 taksa baru—sementara beberapa menyoroti Kosmoceratops sebagai salah satu temuan yang lebih signifikan.[11][12][13][14] Pemerintah AS kemudian digugat oleh sekelompok ilmuwan, aktivis lingkungan, dan penduduk asli Amerika; pada tahun 2021, perluasan monumen ini dipulihkan ke ukuran sebelumnya oleh pemerintahan baru.[10][13][15]
Sebuah tengkorak parsial (dikatalogkan sebagai CMN 8801) yang ditemukan pada tahun 1928 oleh kolektor fosil Charles M. Sternberg di Formasi Dinosaur Park, Alberta, Kanada, ditetapkan sebagai Chasmosaurus russelli pada tahun 1940, namun hanya ditetapkan pada Chasmosaurus pada tahun 1995, karena ketiadaan jumbai leher mencegah tengkorak tersebut diidentifikasi sebagai spesies tertentu. Pada tahun 2014 (dan pada 2015, dalam sebuah artikel yang tidak lulus penelaahan sejawat), ahli paleontologi Nicholas R. Longrich menganggap bahwa tengkorak tersebut mirip dengan Kosmoceratops pada ciri-ciri moncongnya namun berbeda dalam hal bentuk naris dan tanduk hidungnya. Oleh karena itu, ia mengusulkan bahwa fosil tersebut merupakan spesies Kosmoceratops selain K. richardsoni dan menetapkannya sebagai K. sp. (dari spesies yang tidak pasti). Ia beranggapan bahwa masih terlalu dini untuk menamai spesies tersebut karena jumbai leher biasanya diperlukan untuk mendiagnosis suatu spesies ceratopsid, dan sejauh ini baru satu tengkorak yang telah dideskripsikan, sehingga sulit untuk menentukan ciri-ciri dan rentang variasi spesies tersebut.[7][16] Pada tahun 2016, ahli paleontologi James A. Campbell beserta kolega tidak mendukung penempatan spesimen CMN 8801 ke dalam Kosmoceratops, karena mereka mendapati bahwa ciri-ciri yang menjadi dasar penempatannya dipengaruhi oleh tafonomi (perubahan yang terjadi selama pembusukan dan fosilisasi) atau masih berada di dalam rentang variasi antarspesimen Chasmosaurus (meskipun mereka tidak menetapkannya pada spesies tertentu di dalam genus tersebut).[17] Pada tahun 2020, ahli paleontologi Denver W. Fowler dan Elizabeth A. Freedman Fowler menyatakan bahwa penempatan CMN 8801 mungkin dapat dilakukan secara lebih andal saat pemahaman yang lebih baik mengenai anatomi bagian depan tengkorak chasmosaurinae tercapai.[18]
Deskripsi

Kosmoceratops diperkirakan memiliki panjang 45 m (148 ft) dan berat 12 t (13 ton pendek). Sebagai seekor ceratopsid, hewan ini merupakan hewan kuadruped dengan kerangka yang berkonstruksi kokoh, panggul yang besar, dan ekor yang relatif pendek. Hewan ini memiliki barisan gigi utama yang panjang yang membentuk baterai gigi pengiris yang kompleks berisi ratusan gigi di belakang paruh yang edentulosa (tidak bergigi).[19][2] Paruh pramaksila atas dari Kosmoceratops berbentuk segitiga, dan tulang rostral paling depan berbentuk runcing, dengan ujung yang menonjol ke depan dan ke bawah. Pangkal tanduk hidung (tonjolan tulang tempat tumbuhnya tanduk hidung) pada hewan ini tidak biasa di kalangan chasmosaurinae karena bentuknya yang menyerupai bilah, dengan dasar yang memanjang dan menyempit dari sisi ke sisi, serta bagian atas yang memipih. Pangkal tanduk hidung secara keseluruhan berbentuk segi empat dan terletak relatif jauh di belakang pada moncongnya. Naris (bukaan tulang lubang hidung) berbeda dari ceratopsid lainnya karena bentuknya yang tinggi, relatif sempit dari depan ke belakang, dan sangat jelas berbentuk elips (bukan hampir melingkar), dengan kemiringan ke belakang yang menonjol. Penopang narial dari premaksila (yang memanjang dari dasar rongga hidung ke bagian atas) juga miring ke belakang, sebuah ciri yang juga terlihat pada Anchiceratops dan Arrhinoceratops, dan prosesus narial yang menonjol ke belakang dan ke atas dari premaksila berbentuk cabang segitiga.[2][7]

Area di sekitar orbit (rongga mata) sangat khas; sementara sebagian besar chasmosaurinae memiliki pangkal tanduk supraorbital (tanduk di atas mata) yang mengarah ke depan atau ke belakang, Kosmoceratops dan Utahceratops memiliki pangkal tanduk yang mengarah ke atas dan ke samping, secara sekilas mirip dengan bison modern. Dibandingkan dengan pangkal tanduk Utahceratops yang pendek dan tumpul, pangkal tanduk Kosmoceratops lebih memanjang dan ramping, melengkung ke atas kemudian ke bawah, berujung runcing. Tepat di depan dan di tengah orbit, atap tengkoraknya memiliki punuk yang menonjol, yang mana fitur ini hanya ada pada sedikit ceratopsid lain, seperti Diabloceratops. Seperti yang lazim ditemukan pada chasmosaurinae selain Chasmosaurus, dinosaurus ini memiliki osifikasi epijugal ("tanduk pipi") yang relatif besar. Fenestra temporal lateral (bukaan di sisi-sisi tengkorak) berukuran tinggi dan sempit.[2][7]
Jumbai leher parietosquamosal (yang dibentuk oleh tulang parietal dan skuamosal) dari Kosmoceratops sangat terturunkan (atau "maju"), hanya berbagi beberapa ciri dengan Vagaceratops. Hal ini mencakup jumbai yang pendek dari depan ke belakang, dengan fenestra parietal (bukaan yang menembus jumbai) kecil yang terletak jauh di belakang, dan keberadaan sepuluh epiosifikasi menyerupai kail (osifikasi tambahan yang membentuk tanduk dan membingkai tepi jumbai leher pada ceratopsid) di tepi belakang jumbainya. Jumbai Kosmoceratops lebih ekstrem daripada milik Vagaceratops; lebarnya sekitar dua kali dari panjangnya (diukur melintasi permukaan tulang), dengan fenestra parietal yang jauh lebih kecil dan posisinya lebih ke belakang, serta memiliki epiosifikasi yang lebih memanjang dan berbeda dengan jelas di tepi belakangnya. Meskipun secara tradisional chasmosaurinae dianggap sebagai ceratopsid "berjumbai panjang", Kosmoceratops memiliki jumbai paling pendek relatif terhadap lebarnya di antara chasmosaurinae lain dan fenestra parietal terkecil relatif terhadap total area jumbai dibandingkan ceratopsid mana pun. Tulang skuamosal Kosmoceratops melengkung tajam jika dilihat dari samping, dan tepi sampingnya sejajar jika dilihat dari atas. Tulang parietalnya tidak menonjol melewati tulang skuamosal dan memiliki tepi belakang berbentuk V, dengan emarginasi (serangkaian takik) yang membentang di seluruh lebarnya.[2][7]

Epiosifikasi pada tulang skuamosal Kosmoceratops menjadi semakin besar ke arah bagian belakang jumbai leher. Terdapat lima epiosifikasi yang berkembang dengan baik per sisi pada tepi belakang jumbai: tiga epiparietal (ep 1–3) yang melengkung ke depan pada tulang parietal, satu epiparietoskuamosal (esp) yang melengkung ke depan pada perbatasan antara tulang parietal dan skuamosal, dan satu episkuamosal (es1) pada tulang skuamosal yang mengarah ke samping dan ke bawah. Epiparietal yang melengkung ke depan memiliki sulkus (alur) yang menonjol, dan bagian dasarnya menyatu. Dengan lima belas tanduk dan struktur menyerupai tanduk yang berkembang dengan baik, Kosmoceratops memiliki tengkorak paling berhias dari semua dinosaurus yang pernah diketahui; ini mencakup satu pangkal tanduk hidung, dua pangkal tanduk pascaorbital, dua epijugal, dan sepuluh epiosifikasi yang berkembang dengan baik di bagian belakang jumbainya.[2][7] Spesimen subdewasa UMNH VP 16878 memiliki jumlah dan pola epiosifikasi yang sama dengan holotipe dewasa, sehingga memungkinkan untuk membedakan tahap pertumbuhan subdewasa Kosmoceratops dengan tahap pertumbuhan Utahceratops.[3]
Sistem penomoran epiosifikasi yang disajikan di atas mengikuti sistem yang digunakan oleh Sampson beserta kolega pada tahun 2010, namun perlu dicatat bahwa Fowler dan Fowler mengusulkan sistem alternatif pada tahun 2020, berdasarkan perbandingan dengan spesies Chasmosaurus yang lebih awal. Mengikuti sistem ini, epiparietal 1-3 dari Kosmoceratops secara kolektif merupakan ep1 tunggal yang meluas ke samping, epiparietoskuamosal menjadi epiparietal 2, dan episkuamosal menjadi epiparietal 3. Nomor epiosifikasi Vagaceratops juga direvisi dengan cara yang serupa.[2][20]
Klasifikasi

Dua klade (kelompok yang terdiri dari semua taksa yang berbagi nenek moyang yang sama) dinosaurus ceratopsid—Centrosaurinae dan Chasmosaurinae—diakui terutama berdasarkan ornamentasi rumit pada atap tengkoraknya. Pada tahun 2010, Sampson beserta kolega menempatkan Kosmoceratops di kelompok kedua (Chasmosaurinae) karena premaksilanya yang memiliki penopang narial dan prosesus segitiga, serta kehadiran tulang skuamosal yang memanjang. Analisis filogenetik mereka mendapati bahwa Kosmoceratops merupakan takson saudara dari Vagaceratops, di dalam sebuah klade yang dikelompokkan bersama chasmosaurinae terturunkan dari tahap Campanium terbaru dan Maastrichtium pada zaman Kapur Akhir termasuk Triceratops, namun tidak berkerabat dekat dengan Chasmosaurus yang lebih basal (atau "primitif").[2] Pada tahun 2011, ahli paleontologi Jordan C. Mallon dan kolega juga mendapati dukungan untuk klade yang berisi Kosmoceratops dan Vagaceratops, sebagaimana pula ahli paleontologi Steven L. Wick dan Thomas M. Lehman pada tahun 2013.[21][22]
Pada tahun 2011 dan 2014, Longrich mendapati bahwa spesies tunggal Vagaceratops dipertahankan di dalam Chasmosaurus, tempat awalnya ia ditempatkan (sebagai C. irvinensis), sementara Kosmoceratops tidak mengelompok secara dekat dengan taksa lainnya.[23][7] Dalam artikelnya tahun 2015 yang tidak lulus penelaahan sejawat, Longrich berargumen bahwa kemiripan antara jumbai leher keduanya disebabkan oleh evolusi konvergen, yang menyatakan bahwa sepuluh epiosifikasi di bagian belakang jumbai C. irvinensis (atau Vagaceratops, yang mana pada saat itu ia akui bisa jadi merupakan genus yang terpisah) terletak pada tulang parietal tetapi terletak pada tulang parietal dan skuamosal pada Kosmoceratops (Sampson beserta kolega menyatakan bahwa mereka terletak pada parietal dan skuamosal pada keduanya).[16] Dalam sebuah tesis magister tahun 2014, Campbell menyatakan bahwa Sampson beserta kolega awalnya mengidentifikasi kesepuluh epiosifikasi pada tepi belakang jumbai Kosmoceratops dewasa sebagai epiparietal, sebelum menyadari bahwa hanya enam di antaranya yang merupakan epiparietal setelah meneliti spesimen subdewasa, di mana sutura antara tulang parietal dan skuamosal terlihat lebih jelas (sutura menyatu dan menjadi kurang terlihat seiring bertambahnya usia). Oleh karena itu, Sampson beserta kolega juga menafsirkan kembali pola epiosifikasi pada Vagaceratops dengan cara yang sama, namun Campbell mengusulkan bahwa ia malah memiliki empat atau lima epiparietal, sehingga mendapatinya sebagai takson saudara dari Chasmosaurus alih-alih Kosmoceratops.[24] Campbell beserta kolega juga mendapati Vagaceratops berkerabat lebih dekat dengan Chasmosaurus daripada Kosmoceratops pada tahun 2016,[17] dan pada tahun 2019, mereka menganggap V. irvinensis sebagai sebuah spesies dari Chasmosaurus.[25]
Dalam sebuah abstrak konferensi tahun 2011, ahli paleontologi Denver W. Fowler dan kolega mengusulkan bahwa banyak genera ceratopsid hanyalah bentuk ontogenetik (atau tahap pertumbuhan) dari genera lain dan bahwa Kosmoceratops adalah tahap belum matang dari C. irvinensis (dengan ornamentasi tengkorak yang menonjol berkurang seiring bertambahnya usia).[26] Pada tahun 2015, ahli paleontologi Caleb M. Brown dan Donald M. Henderson menguji skema baru untuk homologi di antara epiosifikasi pada chasmosaurinae yang berbeda (yang mana di antaranya saling berkorespondensi antar taksa), dan mendapati klade yang berisi Kosmoceratops dan Vagaceratops sebagai takson saudara dari semua chasmosaurinae lainnya, bertentangan dengan studi-studi sebelumnya.[27] Dalam sebuah buku populer tahun 2016, ahli paleontologi Gregory S. Paul mengusulkan bahwa Kosmoceratops dan Vagaceratops tidak cukup berbeda dari Chasmosaurus untuk menjamin pemisahan genus mereka sendiri dan bahwa spesies mereka harus ditempatkan di dalamnya (atau sebagai alternatif, keduanya harus disatukan di bawah Kosmoceratops), sementara juga mensinonimkan beberapa genera chasmosaurinae lain dengan Chasmosaurus.[19] Sebagian besar genera ceratopsid umumnya dianggap monotipik (berisi spesies tunggal).[28]


Pada tahun 2016, Mallon beserta kolega mendapati Kosmoceratops dan Vagaceratops membentuk sebuah klade, dengan genus baru Spiclypeus sebagai takson saudaranya. Meskipun mereka mengakui bahwa beberapa peneliti lebih mengelompokkan Vagaceratops dengan Chasmosaurus, mereka mendapati epiosifikasi yang menggulung ke depan di bagian belakang jumbai sangatlah khas, menunjukkan bahwa Spiclypeus memberikan petunjuk untuk menjelaskan evolusi fitur ini. Mereka mengusulkan bahwa lengkungan epiosifikasi ini terjadi secara bertahap pada garis keturunan yang mengarah ke Kosmoceratops, epiosifikasi yang berada di tengah menggulung terlebih dahulu, diikuti oleh epiosifikasi di sisi-sisi samping. Proses ini pastinya terjadi sebelum pemendekan jumbai dan penyempitan fenestra parietal, karena fitur-fitur ini berada dalam kondisi yang lebih "primitif" dan tidak berubah pada Spiclypeus. Meskipun Spiclypeus tampaknya bersifat transisional dalam hal ini, ia hidup pada waktu yang bersamaan dengan Kosmoceratops dan oleh karena itu tidak mungkin merupakan nenek moyangnya. Kladogram di bawah ini didasarkan pada analisis Mallon dan kolega pada tahun 2016:[29]
Pada tahun 2020, Fowler dan Fowler mendeskripsikan dua genera chasmosaurinae baru, dan mengusulkan bahwa subfamili tersebut memiliki perpecahan evolusioner yang dalam antara sebuah klade yang berisi Chasmosaurus dan kerabat terdekatnya, serta Pentaceratops dan kerabatnya. Bertentangan dengan studi-studi sebelumnya dan didukung oleh sistem penomoran epiosifikasi baru mereka, Kosmoceratops didapati lebih dekat dengan Chasmosaurus, dan bersama dengan Vagaceratops, mewakili anggota termuda dan paling terturunkan dari garis keturunan tersebut. Menurut interpretasi ini, Kosmoceratops berevolusi dari Vagaceratops, yang mana ia berevolusi dari Chasmosaurus. Karena mereka mendapati Vagaceratops kemungkinan besar merupakan takson saudara dari Kosmoceratops, mereka mengusulkan bahwa ia harus dipertahankan sebagai genus yang terpisah dari Chasmosaurus, karena penempatannya mungkin akan tetap tidak stabil hingga chasmosaurinae lebih dipahami. Spiclypeus tidak dievaluasi dalam studi mereka, di antara chasmosaurinae lain yang baru dinamai.[20]
Paleobiologi

Dalam sebuah tesis magister tahun 2013 (diringkas dalam makalah yang diterbitkan oleh penulis berbeda pada tahun 2019[30]), ahli paleontologi Carolyn Gale Levitt secara histologi meneliti tulang panjang Kosmoceratops (femur dari holotipe dewasa dan spesimen UMNH VP 21339 subdewasa atau dewasa yang ditetapkan padanya) dan Utahceratops untuk memeriksa indikator pertumbuhan dan kedewasaan pada struktur mikro tulang (hingga saat itu satu-satunya chasmosaurinae yang pernah dijadikan sampel untuk hal ini). Jaringan tulang tersebut memiliki jumlah osteosit (sel tulang) yang tinggi serta jaringan pembuluh darah yang padat, termasuk saluran vaskular yang berorientasi radial (saluran darah yang mengalir ke bagian dalam tulang), yang mengindikasikan pertumbuhan cepat yang berkelanjutan. Fitur-fitur ini juga mengindikasikan bahwa ceratopsia memiliki metabolisme yang tinggi dan merupakan hewan endoterm yang homoiotermik (atau "berdarah panas"), seperti burung dan mamalia modern. Tulang Kosmoceratops dan Utahceratops yang dijadikan sampel oleh Levitt tidak menunjukkan bukti adanya garis henti pertumbuhan (garis pertumbuhan tahunan), dan dibandingkan dengan ceratopsid Pachyrhinosaurus, Centrosaurus, dan Einosaurus dari wilayah yang lebih ke utara yang memiliki garis pertumbuhan, hal ini mungkin mengindikasikan bahwa pertumbuhan tulang bereaksi terhadap iklim dan bahwa Kosmoceratops dan Utahceratops dapat mempertahankan pertumbuhan mereka sepanjang tahun karena iklim selatan mereka yang lebih stabil. Ketiadaan garis pertumbuhan juga berarti bahwa usia individu dari spesimen Kosmoceratops dan Utahceratops tidak dapat diperkirakan, namun Levitt menentukan bahwa spesimen Kosmoceratops yang diteliti adalah spesimen subdewasa hingga dewasa, yang menepis klaim Fowler beserta kolega pada tahun 2011 bahwa spesimen tersebut mewakili Vagaceratops yang belum matang. Ia juga menentukan bahwa spesimen Kosmoceratops dan Utahceratops terbesar adalah spesimen dewasa, dan oleh karena itu menepis anggapan bahwa salah satunya merupakan bentuk remaja dari yang lain.[8]
Dalam sebuah tesis magister tahun 2017, ahli paleontologi Nicole Marie Ridgwell mendeskripsikan dua koprolit (kotoran fosil) dari Formasi Kaiparowits yang, berdasarkan ukurannya, mungkin dihasilkan oleh anggota dari salah satu dari tiga kelompok dinosaurus herbivora yang diketahui dari formasi tersebut: ceratopsia (termasuk Kosmoceratops), hadrosaurus, atau ankylosaurus (yang paling langka dari ketiga kelompok tersebut). Koprolit tersebut mengandung fragmen kayu angiosperma (yang mengindikasikan pola makan rambanan berkayu); meskipun sebelumnya hanya ada sedikit bukti bahwa dinosaurus mengonsumsi angiosperma, koprolit-koprolit ini menunjukkan bahwa dinosaurus beradaptasi untuk memakannya (angiosperma baru menjadi umum pada masa Kapur Awal, dan berdiversifikasi pada masa Kapur Akhir). Koprolit tersebut juga mengandung jejak cangkang moluska, kutikula artropoda, dan tulang kadal yang mungkin tertelan bersama materi tumbuhan. Mereka ditemukan di dekat koprolit herbivora lain yang mengandung kayu konifer. Ridgwell mengemukakan bahwa anatomi gigi ceratopsia dan hadrosaurus (dengan baterai gigi yang terdiri dari gigi yang terus diganti) beradaptasi untuk memproses tanaman berserat dalam jumlah besar. Perbedaan pola makan yang diwakili oleh koprolit-koprolit tersebut dapat mengindikasikan adanya segregasi relung di antara herbivora pada ekosistem Formasi Kaiparowits, atau bahwa terdapat variasi pola makan secara musiman.[31]
Fungsi ornamentasi tengkorak

Kemungkinan fungsi tanduk dan jumbai ceratopsia telah diperdebatkan, termasuk untuk menangkis predator, pengenalan spesies, dan pengendalian suhu, meskipun hipotesis dominan melibatkan peningkatan keberhasilan reproduksi. Dalam rilis pers tahun 2010 yang mempresentasikan Kosmoceratops, Utahceratops, dan Vagaceratops, Sampson menyatakan bahwa sebagian besar "fitur aneh" ini tidak akan memadai sebagai senjata untuk melawan predator, tetapi akan digunakan untuk mengintimidasi atau melawan saingan sesama jenis dan menarik perhatian individu dari lawan jenis.[1] Pada tahun 2011, ahli paleontologi Kevin Padian dan John R. Horner mengusulkan bahwa "struktur aneh" pada dinosaurus (termasuk tanduk, jumbai, kubah, dan jambul) terutama digunakan untuk pengenalan spesies (untuk membedakan antar spesies simpatrik; spesies berkerabat yang hidup di area yang sama pada waktu yang sama), dan mereka menolak penjelasan lain karena tidak didukung oleh bukti. Mereka mencatat bahwa banyak ceratopsia besar memiliki bukaan di jumbai mereka, membuatnya tidak banyak berguna dalam pertahanan, dan bahwa berbagai macam ukuran serta orientasi tanduk mereka tidak memiliki fungsi yang jelas dalam pertarungan. Mereka juga mengemukakan bahwa hanya ada sedikit bukti mengenai adanya dimorfisme seksual pada ceratopsia.[32] Menanggapi Padian dan Horner pada tahun yang sama, ahli paleontologi Rob J. Knell dan Sampson berargumen bahwa meskipun pengenalan spesies bisa saja menjadi fungsi sekunder dari "struktur aneh", seleksi seksual (digunakan dalam tampilan visual atau pertarungan untuk bersaing memperebutkan pasangan) merupakan penjelasan yang lebih mungkin karena tingginya biaya untuk mengembangkannya dan bagaimana struktur tersebut tampaknya sangat bervariasi di dalam spesies. Mereka juga mengemukakan bahwa ketiadaan dimorfisme seksual tidak menghalangi penggunaan tanduk dalam persaingan memperebutkan pasangan; bovinae (sapi) jantan menggunakan tanduk mereka untuk tujuan ini sementara betina menggunakannya terutama untuk pertahanan dan sebagai sarana seleksi seksual sekunder.[33]

Pada tahun 2013, ahli paleontologi David E. Hone dan Darren Naish mengkritik "hipotesis pengenalan spesies", dengan berargumen bahwa tidak ada hewan masa kini yang menggunakan struktur semacam itu terutama untuk pengenalan spesies dan bahwa Padian serta Horner telah mengabaikan kemungkinan terjadinya seleksi seksual timbal balik (di mana kedua jenis kelamin memiliki ornamen). Mereka mencatat bahwa jika tujuan utama struktur tersebut adalah pengenalan spesies, mereka akan berbeda dalam bentuk struktur tunggal karena struktur tambahan akan berlebihan dan membutuhkan biaya tambahan. Ceratopsia, misalnya, memiliki tanduk hidung yang rumit, tanduk alis, bos jugal, garis tengah jumbai, dan fitur tepi jumbai, serta perbedaan dalam ukuran dan proporsi tubuh, sementara ketiadaan atau keberadaan tanduk tunggal sudah cukup untuk membedakan antar spesies simpatrik.[34] Pada tahun 2018, ahli paleontologi Andrew Knapp dan kolega meneliti apakah ciri-ciri ornamen ceratopsia yang menyimpang berkorelasi dengan kesimpatrikan antara dua spesies atau lebih, seperti yang diprediksi oleh "hipotesis pengenalan spesies". Mereka mendapati bahwa pola tampilan visual menyimpang secara luas secara keseluruhan sementara pola tampilan spesies simpatrik tidak berbeda secara signifikan dari spesies non-simpatrik, dan menyimpulkan bahwa hipotesis tersebut tidak memiliki dukungan statistik di antara ceratopsia.[35]
Pada tahun 2015, ahli biologi Pasquale Raia beserta kolega meneliti peningkatan evolusioner dalam kerumitan dan ukuran ornamen hewan (seperti jambul, tanduk, dan gading) dari waktu ke waktu, dengan menggunakan amonit, rusa, dan ceratopsia sebagai contoh. Kerumitan jumbai pada ceratopsia tampaknya telah meningkat pada spesies yang lebih baru, dan Kosmoceratops memiliki nilai dimensi fraktal tertinggi di tepi jumbai lehernya (diikuti oleh Styracosaurus, Diabloceratops, dan Centrosaurus). Para penulis mendapati bahwa kerumitan ornamen meningkat seiring dengan ukuran tubuh, yang menunjukkan bahwa evolusi kerumitan ornamen merupakan efek samping dari Aturan Cope (yang mendalilkan bahwa garis keturunan populasi cenderung mengalami peningkatan ukuran tubuh seiring waktu evolusi). Mereka berargumen bahwa meskipun ukuran sebagian besar ornamen mungkin dikendalikan oleh seleksi seksual (dan apakah ornamen tersebut digunakan untuk memilih pasangan, persaingan, atau keduanya), hal tersebut tidak memengaruhi kerumitan dan bentuknya, dan menyimpulkan bahwa kemungkinan besar ukuran alih-alih bentuk ornamenlah yang dipengaruhi oleh seleksi seksual.[36]
Paleolingkungan

Kosmoceratops diketahui dari Formasi Kaiparowits di Utah, yang berasal dari akhir zaman Campanium pada kala Kapur Akhir, dan terdapat dalam rentang stratigrafis yang membentang dari bagian atas dari unit bawahnya hingga bagian atas dari unit tengahnya, pada sedimen yang berasal dari 76,4–75,5 juta tahun yang lalu. Formasi ini diendapkan di bagian selatan sebuah cekungan (Cekungan Pedalaman Barat) di tepi timur daratan yang dikenal sebagai Laramidia (sebuah benua pulau yang terdiri dari wilayah yang sekarang menjadi Amerika Utara bagian barat) dalam jarak 100 km (62 mi) dari Lajur Laut Interior Barat, sebuah laut dangkal di tengah Amerika Utara yang membelah benua tersebut (daratan bagian timurnya dikenal sebagai Appalachia).[2][37][38] Cekungan ini luas, datar, berbentuk bulan sabit, dan dibatasi oleh pegunungan di semua sisi kecuali Lajur Laut Interior Barat di sebelah timur.[39] Formasi ini mewakili lingkungan dataran aluvial hingga pesisir yang basah, lembap, dan didominasi oleh saluran-saluran air yang besar dan dalam dengan tepian yang stabil serta rawa lahan basah, kolam, dan danau abadi. Sungai-sungai umumnya mengalir ke barat melintasi dataran tersebut dan bermuara di Lajur Laut Interior Barat; Pesisir Teluk Amerika Serikat telah diusulkan sebagai analogi modern yang sesuai (seperti rawa Louisiana saat ini). Formasi ini mengawetkan beragam fosil yang melimpah, termasuk hewan darat dan akuatik, tumbuhan, dan palinomorf (mikrofosil organik).[40][6]
Dinosaurus ornithischia lain dari Formasi Kaiparowits mencakup ceratopsia seperti chasmosaurinae Utahceratops, centrosaurinae Nasutoceratops (dan kemungkinan centrosaurinae kedua yang belum dinamai), pachycephalosaurus tak tentu, ankylosaurid Akainacephalus, seekor nodosaurid tak tentu, hadrosaurus Gryposaurus dan Parasaurolophus, serta seekor neornithischia basal tak tentu. Dinosaurus theropoda mencakup tyrannosaurid Teratophoneus, oviraptorosaurus Hagryphus, seekor ornithomimid yang belum dinamai, troodontid Talos, dromaeosaurid tak tentu, dan burung Avisaurus. Hewan vertebrata lainnya mencakup buaya (seperti Deinosuchus dan Brachychampsa), kura-kura (seperti Adocus dan Basilemys), pterosaurus, kadal, ular, amfibi, mamalia, dan ikan.[39][41][42] Dua kelompok vertebrata besar yang paling umum di formasi ini adalah hadrosaurus dan ceratopsia (ceratopsia mewakili sekitar 14 persen dari fosil vertebrata yang berasosiasi dengannya), yang mungkin mengindikasikan kelimpahannya dalam fauna Kaiparowits atau mencerminkan bias pengawetan (sejenis bias pengambilan sampel) karena kelompok-kelompok ini juga memiliki elemen kerangka yang paling kokoh.[4] Telur-telur dari dinosaurus, buaya, dan kura-kura juga telah ditemukan.[43] Rawa-rawa dan lahan basah didominasi oleh pohon sipres setinggi hingga 30 m (98 ft), tumbuhan paku, dan tumbuhan akuatik termasuk mata lele raksasa, apu-apu, serta angiosperma terapung lainnya. Area dengan drainase yang lebih baik didominasi oleh hutan pohon dikotil setinggi hingga 10–20 m (33–66 ft) dan sesekali pohon palem, dengan tumbuhan bawah yang mencakup paku-pakuan. Area dengan drainase baik yang letaknya lebih jauh dari area basah didominasi oleh tumbuhan runjung setinggi hingga 30 m (98 ft), dengan tumbuhan bawah yang terdiri dari sikas, semak atau pohon dikotil kecil, dan kemungkinan tumbuhan paku.[39]

Pada tahun 2010, ahli paleontologi Michael A. Getty beserta kolega meneliti tafonomi holotipe dan spesimen subdewasa UMNH VP 16878 serta keadaan sedimentologi di mana mereka terawetkan. Spesimen holotipe yang kurang lebih terartikulasi ditemukan di litofasies saluran batu pasir berlanau (catatan batuan dari lingkungan sedimen), yang mana sejalan dengan bangkainya yang hanyut ke dalam saluran sungai, kemudian terkubur dengan cepat. Tungkai dan sebagian ekornya mungkin telah hilang karena pemakan bangkai atau pembusukan sebelum bangkai tersebut diendapkan. Bagian-bagian yang hilang dari sisi kiri tengkorak menghilang akibat erosi sebelum penemuan tersebut. Karena sebagian besar kerangka masih dalam tahap preparasi pada saat itu, para peneliti tidak dapat memeriksanya untuk mencari tanda-tanda predasi maupun pemakanan bangkai. Spesimen subdewasa UMNH VP 16878 ditemukan tersebar di area seluas 3 m2 (32 sq ft), dan tingginya tingkat disartikulasinya beserta bagian-bagian yang patah mengindikasikan bahwa spesimen tersebut telah menjadi kerangka dan membusuk sebelum terkubur dalam litofasies batu lumpur berlanau.[4][2] Levitt melaporkan bahwa setiap tulang dari spesimen subdewasa atau dewasa UMNH VP 21339 yang ditetapkan tampak telah patah sebelum diendapkan, dan matriksnya adalah tumpukan batu lanau dan batu lumpur dengan sedikit batu pasir, yang mengindikasikan lingkungan kolam.[8]
Paleobiogeografi

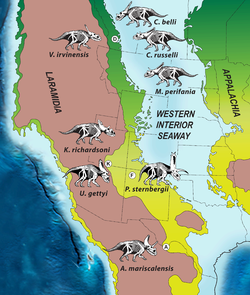
Meskipun luas Laramidia hanya 20 persen dari luas Amerika Utara modern, wilayah ini mengalami radiasi evolusioner dinosaurus yang besar, termasuk hadrosaurus dan ceratopsia yang umum. Telah didalilkan bahwa terdapat susunan latitudinal (berdasarkan garis lintang) "provinsi" atau bioma dinosaurus di Laramidia selama zaman Campanium dan Maastrichtium pada kala Kapur Akhir, dengan batas yang terletak di sekitar Utah utara dan Colorado modern; klade-klade besar yang sama diketahui dari utara dan selatan namun berbeda satu sama lain pada tingkat genus dan spesies. Hipotesis ini telah ditentang; satu argumen mengklaim bahwa kumpulan dinosaurus utara dan selatan selama kurun waktu ini tidaklah hidup sezaman melainkan mencerminkan persebaran taksonomi seiring berjalannya waktu, yang memberikan ilusi adanya provinsi-provinsi yang terisolasi secara geografis, dan bahwa kumpulan yang berbeda tersebut mungkin merupakan artifak dari bias pengambilan sampel di antara formasi-formasi geologi. Akibat kurangnya fosil dengan penanggalan yang baik dari Laramidia selatan, gagasan ini sulit untuk diuji, namun penemuan-penemuan di Formasi Kaiparowits telah meningkatkan pengetahuan tentang vertebrata fosil dari wilayah tersebut selama kala Kapur Akhir. Radiasi evolusioner ceratopsid tampaknya terbatas baik dalam hal waktu maupun geografi (pergantian spesiesnya tinggi, dan masing-masing eksis selama kurang dari satu juta tahun), sebagian besar taksa diketahui dari sedimen Kapur terbaru di Cekungan Pedalaman Barat, sehingga tampaknya berasal dan berdiversifikasi di Laramidia.[2][44]
Penemuan Kosmoceratops dan Utahceratops sangat meningkatkan jumlah chasmosaurinae yang diketahui dari Cekungan Pedalaman Barat. Sampson beserta kolega memandang hal ini sebagai dukungan terkuat untuk gagasan "provinsi" dinosaurus dalam deskripsi mereka pada tahun 2010. Mereka mengemukakan bahwa berbeda dengan Maastrichtium, tahap Campanium sebelumnya memiliki kumpulan dinosaurus yang sampelnya diambil dengan lebih baik, beragam, dan tersebar luas, serta data geografis dan stratigrafis yang lebih presisi. Rentang stratigrafis Kosmoceratops dan Utahceratops menunjukkan bahwa mereka hidup pada waktu yang sama dan kemungkinan besar di ekosistem yang sama, yang mana ini merupakan hal langka di antara ceratopsid. Menurut Sampson dan kolega, penanggalan geokronologis mengindikasikan bahwa interval geologis singkat yang terawetkan di dalam Formasi Kaiparowits sezaman dengan Formasi Dinosaur Park di Alberta dan mengikuti formasi-formasi lain di utara (formasi Judith River atas dan Two Medicine) serta tenggara (formasi Fruitland dan Aguja), dengan durasi singkat, tumpang tindih waktu, dan rentang bujur yang memungkinkan perbandingan signifikan di antara formasi-formasi tersebut. Fakta bahwa Kosmoceratops dan Utahceratops tidak berkerabat dekat satu sama lain maupun dengan Chasmosaurus dan Mojoceratops dari Formasi Dinosaur Park yang sezaman, dan bahwa Vagaceratops dari Alberta tumpang tindih secara waktu dengan Pentaceratops dari New Mexico, dikutip oleh Sampson beserta kolega sebagai bukti yang menentang klaim bahwa kumpulan dinosaurus utara dan selatan tidak hidup sezaman selama kurun waktu ini.[2]

Sampson beserta kolega menyatakan bahwa studi mereka merupakan pendokumentasian endemisme intrakontinental pada dinosaurus untuk pertama kalinya (dengan taksa chasmosaurinae yang berbeda muncul secara bersamaan di utara dan selatan Laramidia). Mereka mengusulkan urutan peristiwa dalam evolusi chasmosaurinae yang mereka dapati konsisten dengan bukti filogenetik, stratigrafik, dan biogeografik. Menurut mereka, chasmosaurinae berasal dari Laramidia sekitar 80–90 juta tahun yang lalu dan menyebar ke sebagian besar daratan ini pada 77 juta tahun yang lalu. Pada saat itu, sebuah penghalang telah muncul di area Utah dan Colorado, mencegah hewan-hewan menyebar di antara Laramidia utara dan selatan serta menyebabkan chasmosaurinae utara dan selatan (dan mungkin klade hewan lainnya) berevolusi secara independen melalui vikariansi di dalam pusat endemisme terpisah yang tersusun secara latitudinal pada 77,0–75,8 juta tahun yang lalu. Setelah penghalang tersebut lenyap sekitar 75,7 juta tahun yang lalu, garis keturunan Kosmoceratops (diwakili oleh Vagaceratops) yang sebelumnya terbatas di Laramidia selatan menyebar ke utara, dan memunculkan semua chasmosaurinae yang muncul belakangan, seperti Anchiceratops dan Triceratops.[2]
Meskipun dinosaurus Campanium akhir di Laramidia berukuran lebih besar daripada kebanyakan hewan modern bertubuh besar (yang membutuhkan rentang persebaran spesies yang luas akibat tingginya kebutuhan makan), Sampson beserta kolega mendapati bahwa mereka tampaknya memiliki rentang spesies yang relatif kecil, yang mana hal ini semakin membingungkan jika mengingat tingginya keanekaragaman spesies dari dinosaurus Laramidia. Walaupun mereka tampaknya mendiami setidaknya dua wilayah yang semi-terisolasi, tidak ada bukti adanya penghalang penyebaran, dan terdapat gradien suhu yang lebih kecil dibandingkan masa kini. Dinosaurus di sana tampaknya peka terhadap zonasi latitudinal lingkungan (berpotensi karena tingkat fisiologi yang lebih rendah atau lingkungannya yang lebih produktif), yang kemungkinan bertahan selama setidaknya 1,5 juta tahun. Alternatif dari keberadaan penghalang mencakup gagasan bahwa provinsi-provinsi diskret tersebut dipisahkan oleh zona percampuran fauna, atau bahwa terdapat gradien atau klinal yang berkelanjutan di seluruh ketinggian, tanpa zona endemik yang berbeda dengan jelas. Kemungkinan penghalang fisik untuk penyebaran mencakup barisan pegunungan dari timur ke barat yang belum teridentifikasi, banjir di wilayah dataran akibat Lajur Laut Interior Barat (yang akan melenyapkan habitat elevasi rendah di Laramidia tengah untuk sementara waktu), atau sistem sungai besar. Sampson beserta kolega menganggap lebih mungkin bahwa terdapat penghalang paleoklimatik atau paleolingkungan terhadap penyebaran (sebuah gagasan yang didukung oleh jenis serbuk sari yang berbeda di Laramidia utara dan selatan), namun mencatat bahwa lebih banyak bukti diperlukan untuk menyelidiki sifat pemisahan antara provinsi-provinsi fauna di Laramidia.[2]
Pada tahun 2012, ahli paleontologi Terry A. Gates beserta kolega mengusulkan bahwa peningkatan keanekaragaman hayati dinosaurus Amerika Utara selama zaman Campanium disebabkan oleh peristiwa orogenik (yang berujung pada perubahan di kerak Bumi tempat bertemunya lempeng benua) di Cekungan Pedalaman Barat, termasuk awal pertemuan Sabuk Orogenik Sevier dan lempeng tektonik di Laramidia, yang membentuk pegunungan yang mengisolasi ceratopsid dan hadrosaurid serta mendorong diversifikasi mereka. Mereka mengusulkan bahwa perpisahan Kosmoceratops dan Vagaceratops dari klade yang mencakup Anchiceratops dan Triceratops disebabkan oleh peristiwa vikariansi ataupun penyebaran.[45] Setelah menyimpulkan pada tahun 2014 bahwa Vagaceratops berkerabat lebih dekat dengan Chasmosaurus daripada Kosmoceratops, Campbell mengusulkan bahwa Vagaceratops berasal dari Laramidia utara. Ia membantah bahwa hewan tersebut telah bermigrasi ke sana dari selatan, seperti yang diklaim oleh Sampson beserta kolega (sehingga menjadikan namanya, "wajah bertanduk yang mengembara", sebagai sebuah salah kaprah).[24][1]

Melalui penetapan tengkorak CMN 8801 pada Kosmoceratops sp. pada tahun 2014 serta penamaan spesies baru Pentaceratops (P. aquilonius) olehnya, yang keduanya berasal dari Dinosaur Park di Alberta, Longrich berargumen menentang gagasan adanya provinsi dinosaurus utara dan selatan yang terpisah, mengingat kedua genera tersebut sekarang diketahui baik dari Amerika Utara bagian tenggara maupun utara. Mengingat hewan-hewan utara memang berbeda dari rekan-rekannya di selatan, ia sepakat bahwa terdapat endemisme yang signifikan selama zaman Campanium, namun mendapati bahwa garis keturunan dinosaurus mampu menyebar melintasi jarak yang jauh dan tidak terkendala oleh penghalang geografis, iklim, atau flora, dan endemisme alih-alih merupakan hasil dari populasi yang telah mapan (serta beradaptasi secara lokal) dalam menyingkirkan dinosaurus pendatang melalui persaingan.[7] Pada tahun 2016, Campbell beserta kolega menetapkan CMN 8801 kembali pada Chasmosaurus, dan mendukung gagasan bahwa Kosmoceratops terbatas persebarannya di Laramidia selatan.[17] Pada tahun yang sama, Mallon beserta kolega mendapati bahwa P. aquilonius merupakan nomen dubium (nama yang meragukan) yang tidak valid dan sepakat bahwa terdapat penghalang penyebaran di antara Laramidia utara dan selatan. Mereka mengusulkan bahwa hubungan kekerabatan yang dekat antara Kosmoceratops dan Spiclypeus dari Montana di wilayah utara mengindikasikan pernah adanya koneksi geografis yang berusia pendek.[29]
Pada tahun 2016, ahli paleontologi Spencer G. Lucas beserta kolega mengkritik gagasan provinsialisme dinosaurus utara–selatan di Laramidia, dan mendapati bukti yang mendukung endemisme di antara vertebrata non-dinosaurus lemah, dan pada dinosaurus bermasalah, karena utamanya hanya segelintir taksa chasmosaurinae yang digunakan untuk mengidentifikasinya. Mereka menolak gagasan mengenai endemisme dan provinsialisme dinosaurus akibat berbagai masalah terkait bias pengambilan sampel (kesan bahwa dinosaurus berdiversifikasi selama zaman Campanium merupakan hasil dari catatan fosil yang lebih padat dari masa tersebut), ketiadaan penghalang topografik yang akan membagi provinsi-provinsi tersebut (Gates beserta kolega disinyalir telah menyalahartikan efek topografik orogeni di Laramidia), kurangnya perbedaan iklim maupun vegetasi yang signifikan, keputusan-keputusan taksonomi yang telah dilibatkan dalam mempersepsikan gagasan ini, diakroneitas (perbedaan usia) sebagian besar kumpulan fosil yang mencegah penggunaannya dalam analisis biogeografik, dan bahwa kesimpulan dari pihak-pihak yang mendukung gagasan ini tidak seragam serta melemahkan argumen mereka sendiri. Sebagai contoh, mereka mengemukakan bahwa bertentangan dengan klaim yang dilontarkan oleh Sampson dan kolega, Kosmoceratops dan Utahceratops tidak sezaman dengan Vagaceratops (yang berusia lebih muda) dan ketiganya berusia lebih tua daripada Pentaceratops. Lucas beserta kolega mengakui bahwa terdapat beberapa endemisme yang tidak dapat disebabkan oleh bias pengambilan sampel, namun mereka sepakat dengan Longrich bahwa hal ini mungkin disebabkan oleh persaingan atau spesialisasi pola makan.[46]

Pada tahun 2017, Fowler menunjukkan bahwa penanggalan Formasi Kaiparowits yang telah digunakan oleh Sampson beserta kolega untuk mendemonstrasikan bahwa Kosmoceratops dan Utahceratops hidup sezaman dengan chasmosaurinae dari formasi-formasi lain di utara, telah dikalibrasi secara berbeda dari penanggalan formasi lainnya. Ketika dikalibrasi ulang dengan standar yang sama, ia mendemonstrasikan bahwa tumpang tindih stratigrafis antara taksa-taksa tersebut tidak dikenali. Sebaliknya, hanya bagian bawah dari Formasi Kaiparowits yang tumpang tindih dengan Formasi Dinosaur Park, namun bagian ini tidak mengandung Kosmoceratops dan Utahceratops, sementara sisa-sisa yang terfragmentasi menunjukkan bahwa beberapa taksa muncul bersama di kedua formasi tersebut pada rentang ini. Oleh karena itu, Fowler merasa lebih mungkin bahwa perbedaan pada taksa dinosaurus di antara formasi-formasi tersebut disebabkan oleh pengambilan sampel tingkat stratigrafis yang berbeda alih-alih segregasi biogeografik, sebuah penjelasan yang juga ia rasa memungkinkan untuk menjelaskan perbedaan antara Formasi Kaiparowits dengan formasi Fruitland dan Kirtland.[38]
Walaupun tidak mendukung adanya endemisme, Fowler dan Fowler mengusulkan pada tahun 2020 bahwa mungkin telah terdapat provinsialisme gradasional yang samar pada chasmoraurinae dari Utara ke Selatan, dengan garis keturunan yang mencakup Chasmosaurus lebih melimpah di Utara, Kosmoceratops menjadi anggota paling selatannya, sementara garis keturunan yang mencakup Pentaceratops lebih melimpah di Selatan. Mengingat kedua klade tersebut tumpang tindih secara geografis selama bagian paling atas dari Campanium tengah, peristiwa spesiasi yang berujung pada kedua garis keturunan tersebut mungkin disebabkan oleh vikariansi latitudinal sebelum munculnya anggota pertama garis keturunan Chasmosaurus, 77 juta tahun yang lalu. Pada 76 juta tahun yang lalu, dugaan penghalang yang memisahkan garis keturunan tersebut pastinya telah lenyap, karena mereka hidup berdampingan setelahnya; garis pemisahnya tampaknya berlokasi di suatu tempat di antara Utah selatan dan Montana utara.[20] Fowler dan Fowler juga mendapati bahwa Vagaceratops hidup sekitar 76,2 juta tahun yang lalu sementara Kosmoceratops hidup sekitar 76–75,9 juta tahun yang lalu, dan tidak tumpang tindih secara stratigrafis.[18]
Lihat pula
Referensi
- ^ a b c d e f "Amazing horned dinosaurs unearthed on "lost continent"". University of Utah News. 2010. Diarsipkan dari asli tanggal Februari 22, 2011. Diakses tanggal Juli 3, 2019.
- ^ a b c d e f g h i j k l m n o p q r s t Sampson, S. D.; Loewen, M. A.; Farke, A. A.; Roberts, E. M.; Forster, C. A.; Smith, J. A.; Titus, A. L.; Stepanova, A. (2010). "New horned dinosaurs from Utah provide evidence for intracontinental dinosaur endemism". PLoS ONE. 5 (9) e12292. Bibcode:2010PLoSO...512292S. doi:10.1371/journal.pone.0012292. PMC 2929175. PMID 20877459.
- ^ a b c d e f Loewen, M.; Farke, A. A.; Sampson, S. D.; Getty, M. A.; Lund, E. K.; O'Connor, P. M. (2013). "Ceratopsid dinosaurs from the Grand Staircase of Southern Utah". Dalam Titus, A.; Loewen, M. (ed.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. hlm. 488–503. ISBN 978-0-253-00883-1.
- ^ a b c d Getty, M. A.; Loewen, M. A.; Roberts, E.; Titus, A. L.; Sampson, S. D. (2010), "Taphonomy of horned dinosaurs (Ornithischia: Ceratopsidae) from the late Campanian Kaiparowits Formation, Grand Staircase – Escalante National Monument, Utah", dalam Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A. (ed.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Bloomington: Indiana University Press, hlm. 478–494, ISBN 978-0-253-35358-0
- ^ Lund, Eric K.; Sampson, Scott D.; Loewen, Mark A. (2016). "Nasutoceratops titusi (Ornithischia, Ceratopsidae), a basal centrosaurine ceratopsid from the Kaiparowits Formation, southern Utah". Journal of Vertebrate Paleontology. 36 (2) e1054936. doi:10.1080/02724634.2015.1054936.
- ^ a b Sampson, S. D. (2012). "Dinosaurs of the lost continent". Scientific American. 306 (3): 40–47. Bibcode:2012SciAm.306c..40S. doi:10.1038/scientificamerican0312-40. ISSN 0036-8733. JSTOR 26014255. PMID 22375321.
- ^ a b c d e f g h Longrich, N. R. (2014). "The horned dinosaurs Pentaceratops and Kosmoceratops from the upper Campanian of Alberta and implications for dinosaur biogeography". Cretaceous Research. 51: 292–308. doi:10.1016/j.cretres.2014.06.011.
- ^ a b c Levitt, C. G. (2013). Bone histology and growth of chasmosaurine ceratopsid dinosaurs from the late Campanian Kaiparowits Formation, Southern Utah (Thesis). University of Utah. Diarsipkan dari versi aslinya tanggal Juli 14, 2017. Diakses tanggal Juni 25, 2019.
- ^ Dodson, P. (2013). "Ceratopsia increase: history and trends". Canadian Journal of Earth Sciences. 50 (3): 294–305. Bibcode:2013CaJES..50..294D. doi:10.1139/cjes-2012-0085.
- ^ a b Gonzales, R.; Siegler, K.; Dwyer, C. (2017). "Trump orders largest national monument reduction In U.S. history". NPR.org. Diarsipkan dari versi aslinya tanggal Juli 5, 2019. Diakses tanggal Juni 25, 2019.
- ^ a b Finnegan, M. (2017). "Remarkable dinosaur discoveries under threat with Trump plan to shrink national monument in Utah, scientists say". Los Angeles Times. Diarsipkan dari versi aslinya tanggal Juni 25, 2019. Diakses tanggal Juni 24, 2019.
- ^ Panciroli, E. (2017). "Why are palaeontologists suing Trump?". The Guardian. Diarsipkan dari versi aslinya tanggal Mei 31, 2019. Diakses tanggal Juni 25, 2019.
- ^ a b Wei-Haas, M. (2017). "What shrinking fossil-rich national monuments means for science". Smithsonian Magazine. Diarsipkan dari versi aslinya tanggal Juni 24, 2019. Diakses tanggal Juni 24, 2019.
- ^ Gramling, C. (2017). "Science and politics collide over Bears Ears and other national monuments". Science. doi:10.1126/science.aal1122. Diarsipkan dari versi aslinya tanggal Juni 26, 2019. Diakses tanggal Juni 26, 2019.
- ^ "The Wilderness Society et al.v. Trump et al. (Grand Staircase–Escalante)". NRDC. April 10, 2020. Diarsipkan dari versi aslinya tanggal April 2, 2021. Diakses tanggal April 19, 2020.
- ^ a b Longrich, N. R. (2015). "Systematics of Chasmosaurus – new information from the Peabody Museum skull, and the use of phylogenetic analysis for dinosaur alpha taxonomy". F1000Research. 4: 1468. doi:10.12688/f1000research.7573.1.
- ^ a b c Campbell, J. A.; Ryan, M. J.; Holmes, R. B.; Schröder-Adams, C. J.; Mihlbachler, M. C. (2016). "A re-evaluation of the chasmosaurine ceratopsid genus Chasmosaurus (Dinosauria: Ornithischia) from the Upper Cretaceous (Campanian) Dinosaur Park Formation of Western Canada". PLOS ONE. 11 (1) e0145805. Bibcode:2016PLoSO..1145805C. doi:10.1371/journal.pone.0145805. PMC 4699738. PMID 26726769.
- ^ a b Fowler, D. W.; Freedman Fowler, E. A. (2020). "Transitional evolutionary forms in chasmosaurine ceratopsid dinosaurs: evidence from the Campanian of New Mexico". PeerJ. 8: e9251. doi:10.7717/peerj.9251/supp-1. PMID 32547873.
- ^ a b Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs (Edisi 2). Princeton: Princeton University Press. hlm. 285–286, 296. ISBN 978-0-691-16766-4.
- ^ a b c Fowler, D. W.; Freedman Fowler, E. A. (2020). "Transitional evolutionary forms in chasmosaurine ceratopsid dinosaurs: evidence from the Campanian of New Mexico". PeerJ. 8 e9251. doi:10.7717/peerj.9251. PMC 7278894.
- ^ Mallon, J. C.; Holmes, R.; Eberth, D. A.; Ryan, M. J.; Anderson, J. S. (2011). "Variation in the skull of Anchiceratops (Dinosauria, Ceratopsidae) from the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta". Journal of Vertebrate Paleontology. 31 (5): 1047–1071. doi:10.1080/02724634.2011.601484.
- ^ Wick, S. L.; Lehman, T. M. (2013). "A new ceratopsian dinosaur from the Javelina Formation (Maastrichtian) of West Texas and implications for chasmosaurine phylogeny". Naturwissenschaften. 100 (7): 667–682. Bibcode:2013NW....100..667W. doi:10.1007/s00114-013-1063-0. PMID 23728202.
- ^ Longrich, N. R. (2011). "Titanoceratops ouranos, a giant horned dinosaur from the late Campanian of New Mexico". Cretaceous Research. 32 (3): 264–276. doi:10.1016/j.cretres.2010.12.007.
- ^ a b Campbell, J. A. (2014). A re-evaluation of the chasmosaurine ceratopsid Chasmosaurus (Dinosauria: Ornithischia) from the Upper Cretaceous (Campanian) Dinosaur Park Formation of Western Canada (Thesis). Carleton University. doi:10.22215/etd/2014-10978. Diarsipkan dari versi aslinya tanggal Juni 30, 2019. Diakses tanggal Juni 30, 2019.
- ^ Campbell, J. A.; Ryan, M. J.; Schröder-Adams, C. J.; Holmes, R. B.; Evans, D. C. (2019). "Temporal range extension and evolution of the chasmosaurine ceratopsid 'Vagaceratops' irvinensis (Dinosauria: Ornithischia) in the Upper Cretaceous (Campanian) Dinosaur Park Formation of Alberta". Vertebrate Anatomy Morphology Palaeontology. 7: 83–100. doi:10.18435/vamp29356.
- ^ Fowler, D. W.; Scanella, J.; Horner, J. R. (2011). "Reassessing ceratopsid diversity using unified frames of reference". Journal of Vertebrate Paleontology, SVP Program and Abstracts Book. hlm. 111A.
- Also published as Fowler, D. W.; Scanella, J.; Horner, J. R. (2011). "Reassessing ceratopsid diversity using unified frames of reference". Journal of Vertebrate Paleontology. 31 (Supplement 2): 111. doi:10.1080/02724634.2011.10635174.
- ^ Brown, C. M.; Henderson, D. M. (2015). "A new horned dinosaur reveals convergent evolution in cranial ornamentation in Ceratopsidae". Current Biology. 25 (12): 1641–1648. doi:10.1016/j.cub.2015.04.041. PMID 26051892.
- ^ Lehman, T. M.; Wick, S. L.; Barnes, K. R. (2016). "New specimens of horned dinosaurs from the Aguja Formation of West Texas, and a revision of Agujaceratops". Journal of Systematic Palaeontology. 15 (8): 641–674. doi:10.1080/14772019.2016.1210683.
- ^ a b Mallon, J. C.; Ott, C. J.; Larson, P. L.; Iuliano, E. M.; Evans, D. C.; Evans, A. R. (2016). "Spiclypeus shipporum gen. et sp. nov., a boldly audacious new chasmosaurine Ceratopsid (Dinosauria: Ornithischia) from the Judith River Formation (Upper Cretaceous: Campanian) of Montana, USA". PLOS ONE. 11 (5) e0154218. Bibcode:2016PLoSO..1154218M. doi:10.1371/journal.pone.0154218. PMC 4871577. PMID 27191389.
- ^ Hedrick, B. P.; Goldsmith, E.; Rivera-Sylva, H.; Fiorillo, A. R.; Tumarkin-Deratzian, A. R.; Dodson, P. (2019). "Filling in gaps in the ceratopsid histologic database: histology of two basal centrosaurines and an assessment of the utility of rib histology in the Ceratopsidae". The Anatomical Record: 40. doi:10.1002/ar.24099. PMID 30773832.
- ^ Ridgwell, N. M. (2017). Description of Kaiparowits coprolites that provide rare direct evidence of angiosperm consumption by dinosaurs. Museum and Field Studies Graduate Theses & Dissertations (Thesis). Diarsipkan dari versi aslinya tanggal Juni 30, 2019. Diakses tanggal Juni 30, 2019.
- ^ Padian, K.; Horner, J. R. (2011). "The evolution of 'bizarre structures' in dinosaurs: biomechanics, sexual selection, social selection or species recognition?". Journal of Zoology. 283 (1): 3–17. doi:10.1111/j.1469-7998.2010.00719.x.
- ^ Knell, R. J.; Sampson, S. (2011). "Bizarre structures in dinosaurs: species recognition or sexual selection? A response to Padian and Horner" (PDF). Journal of Zoology. 283 (1): 18–22. doi:10.1111/j.1469-7998.2010.00758.x. Diarsipkan (PDF) dari versi aslinya tanggal Juli 22, 2018. Diakses tanggal Juni 25, 2019.
- ^ Hone, D. W. E.; Naish, D. (2013). "The 'species recognition hypothesis' does not explain the presence and evolution of exaggerated structures in non-avialan dinosaurs". Journal of Zoology. 290 (3): 172–180. doi:10.1111/jzo.12035.
- ^ Knapp, A.; Knell, R. J.; Farke, A. A.; Loewen, M. A.; Hone, D. W. E. (2018). "Patterns of divergence in the morphology of ceratopsian dinosaurs: sympatry is not a driver of ornament evolution". Proceedings of the Royal Society B: Biological Sciences. 285 (1875) 20180312. doi:10.1098/rspb.2018.0312. PMC 5897650. PMID 29563271.
- ^ Raia, P.; Passaro, F.; Carotenuto, F.; Maiorino, L.; Piras, P.; Teresi, L.; Meiri, S.; Itescu, Y.; Novosolov, M.; Baiano, M. A.; Martinez, R.; Fortelius, M. (2015). "Cope's rule and the universal scaling law of ornament complexity". The American Naturalist. 186 (2): 165–175. doi:10.1086/682011. hdl:11336/55178. PMID 26655146.
- ^ Roberts, E. M.; Deino, A. L.; Chan, M. A. (2005). "40Ar/39Ar age of the Kaiparowits Formation, southern Utah, and correlation of contemporaneous Campanian strata and vertebrate faunas along the margin of the Western Interior Basin". Cretaceous Research. 26 (2): 307–318. doi:10.1016/j.cretres.2005.01.002.
- ^ a b Fowler, D. W.; Wong, William O. (2017). "Revised geochronology, correlation, and dinosaur stratigraphic ranges of the Santonian-Maastrichtian (Late Cretaceous) formations of the Western Interior of North America". PLOS ONE. 12 (11) e0188426. Bibcode:2017PLoSO..1288426F. doi:10.1371/journal.pone.0188426. PMC 5699823. PMID 29166406.
- ^ a b c Sampson, S. D.; Loewen, M. A.; Roberts, E. M.; Getty, M. A. (2013). "A new macrovertebrate assemblage from the Late Cretaceous (Campanian) of Southern Utah". Dalam Titus, A.; Loewen, M. (ed.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. hlm. 599–622. ISBN 978-0-253-00883-1.
- ^ Roberts, E. M.; Sampson, S. D.; Deino, A. L.; Bowring, S. A.; Buchwaldt, S. (2013). "The Kaiparowits Formation: a remarkable record of Late Cretaceous terrestrial environments, ecosystems, and evolution in Western North America". Dalam Titus, A.; Loewen, M. (ed.). At the Top of the Grand Staircase: The Late Cretaceous of Southern Utah. Bloomington: Indiana University Press. hlm. 85–106. ISBN 978-0-253-00883-1.
- ^ Titus, A. L.; Eaton, J. G.; Sertich, J. (2016). "Late Cretaceous stratigraphy and vertebrate faunas of the Markagunt, Paunsaugunt, and Kaiparowits plateaus, southern Utah". Geology of the Intermountain West. 3: 229–291. doi:10.31711/giw.v3i0.10.
- ^ Wiersma, J. P.; Irmis, R. B. (2018). "A new southern Laramidian ankylosaurid, Akainacephalus johnsoni gen. et sp. nov., from the upper Campanian Kaiparowits Formation of southern Utah, USA". PeerJ. 6: 76. doi:10.7717/peerj.5016. PMC 6063217. PMID 30065856.
- ^ Oser, S. E. (2018). Campanian ooassemblages within the Western Interior Basin: eggshell from the Upper Cretaceous Kaiparowits Formation of Grand Staircase–Escalante National Monument, UT (MSc thesis). Boulder: Department of Museum and Field Studies, University of Colorado. Diarsipkan dari versi aslinya tanggal Juli 5, 2019. Diakses tanggal Juli 5, 2019.
- ^ Sampson, S. D.; Loewen, M. A. (2010), "Unraveling a radiation: a review of the diversity, stratigraphic distribution, biogeography, and evolution of horned dinosaurs. (Ornithischia: Ceratopsidae)", dalam Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A. (ed.), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Bloomington: Indiana University Press, hlm. 405–427, ISBN 978-0-253-35358-0
- ^ Gates, T. A.; Prieto-Márquez, A.; Zanno, L. E.; Dodson, P. (2012). "Mountain building triggered Late Cretaceous North American megaherbivore dinosaur radiation". PLoS ONE. 7 (8) e42135. Bibcode:2012PLoSO...742135G. doi:10.1371/journal.pone.0042135. PMC 3410882. PMID 22876302.
- ^ Lucas, S. G.; Sullivan, R. M.; Lichtig, A.; Dalman, S. (2016). "Late Cretaceous dinosaur biogeography and endemism in the Western Interior basin, North America: A critical re-evaluation". New Mexico Museum of Natural History and Science Bulletin. 71: 195–213.
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||




Konten ini disalin dari wikipedia, mohon digunakan dengan bijak.

