Mata
| Mata | |
|---|---|
 Mata manusia | |
 Mata majemuk kril antartika | |
| Rincian | |
| Sistem | Saraf |
| Pengidentifikasi | |
| Bahasa Latin | oculus |
| TA98 | A15.2.00.001 A01.1.00.007 |
| TA2 | 113, 6734 |
| Daftar istilah anatomi | |
Mata,[1] atau netra[2] adalah organ penglihatan. Mata mendeteksi cahaya dan mengubahnya menjadi impuls elektrokimia pada sel saraf. Pada organisme yang lebih tinggi, mata adalah sistem optik kompleks yang mengumpulkan cahaya dari lingkungan sekitarnya, mengatur intensitasnya melalui diafragma, memfokuskan melalui penyesuaian lensa untuk membentuk sebuah gambar, mengkonversi gambar tersebut menjadi satu himpunan sinyal listrik, dan mentransmisikan sinyal-sinyal ke otak melalui jalur saraf kompleks yang menghubungkan mata melalui saraf optik menuju korteks visual dan area lain dari otak. Mata dengan daya resolusinya memiliki sepuluh bentuk yang berbeda secara fundamental, dan 96% dari spesies hewan memiliki sistem optik yang kompleks.[3] Mata kompleks ini mampu membentuk gambar, seperti pada moluska, kordata, dan artropoda.[4]
"Mata" yang paling sederhana, seperti pada mikroorganisme, tidak melakukan apa-apa, tetapi dapat mendeteksi apakah lingkungan sekitarnya terang atau gelap, yang cukup untuk pengiringarusan ritme sirkadian.[5] Dari mata yang lebih kompleks, sel ganglion fotosensitif retina mengirim sinyal sepanjang saluran retinohipotalamik menuju inti suprakiasmatik untuk efek penyesuaian sirkadian dan menuju daerah pratektal untuk mengontrol refleks sinar pupilar.
Tinjauan luas


Mata kompleks dapat membedakan bentuk dan warna. Bidang penglihatan pada banyak organisme, terutama predator, penglihatan binokular melibatkan wilayah luas untuk meningkatkan persepsi kedalaman. Pada organisme lain, mata terletak sedemikian rupa sehingga memaksimalkan bidang pandang, seperti pada kelinci dan kuda, yang memiliki penglihatan monokular.
Proto-mata pertama di antara hewan berevolusi 600 juta tahun silam sekitar ledakan Kambrium.[6] Nenek moyang terakhir dari hewan memiliki perangkat biokimia yang diperlukan untuk penglihatan, dan mata lebih maju berkembang pada 96% dari spesies hewan pada enam dari ~35[a] filum utama.[3] Pada kebanyakan vertebrata dan beberapa moluska, mata bekerja dengan memungkinkan cahaya untuk masuk dan memproyeksikannya pada panel sel peka cahaya, yang dikenal sebagai retina, di belakang mata. Sel kerucut (untuk warna) dan sel batang (untuk kontras cahaya rendah) pada retina mendeteksi dan mengkonversi cahaya menjadi sinyal saraf untuk penglihatan. Sinyal visual tersebut kemudian diteruskan ke otak melalui saraf optik. Mata biasanya berbentuk seperti bola, diisi dengan zat seperti gel transparan yang disebut badan bening, dengan lensa pemfokus dan sering dengan suatu selaput pelangi; relaksasi atau kontraksi otot-otot di sekitar selaput pelangi mengubah ukuran pupil, sehingga mengatur jumlah cahaya yang masuk ke mata,[7] dan mengurangi aberasi ketika terdapat cahaya yang cukup.[8] Mata pada kebanyakan sefalopoda, ikan, amfibi, dan ular telah memiliki bentuk lensa yang tetap, dan memfokuskan penglihatan yang diperoleh dengan lensa teleskopik—serupa dengan cara kamera berfokus.[9]
Mata majemuk ditemukan pada artropoda dan terdiri dari banyak faset sederhana yang, tergantung pada detail anatomi, dapat memberikan baik citra terpikselasi tunggal maupun beberapa gambar per mata. Setiap sensor memiliki lensa sendiri dan sel fotosensitif. Beberapa mata memiliki hingga 28.000 sensor tersebut, yang diatur secara heksagonal, dan dapat memberikan bidang penglihatan 360° penuh. Mata majemuk sangat sensitif terhadap gerakan. Beberapa artropoda, termasuk banyak Strepsiptera, memiliki mata majemuk dari hanya beberapa faset, masing-masing dengan retina yang mampu membuat gambar, menciptakan penglihatan. Dengan setiap mata melihat sesuatu yang berbeda, gambar menyatu dari semua mata dan dihasilkannya gambar yang sangat berbeda dan beresolusi tinggi di dalam otak.
Memiliki penglihatan warna hiperspektral mendetail, udang sentadu telah dilaporkan memiliki sistem penglihatan warna paling kompleks di dunia.[10] Trilobita, yang sekarang sudah punah, memiliki mata majemuk yang unik. Hewan ini menggunakan kristal kalsit bening untuk membentuk lensa mata. Dalam hal ini, Trilobita berbeda dari kebanyakan artropoda lainnya yang memiliki mata lembut. Jumlah lensa mata bervariasi, tetapi beberapa trilobita hanya memiliki satu lensa, dan beberapa memiliki ribuan lensa dalam satu mata.
Berbeda dengan mata majemuk, mata sederhana adalah mata yang memiliki lensa tunggal. Misalnya, laba-laba peloncat memiliki sepasang mata sederhana yang besar dengan ruang pandang sempit, didukung oleh susunan lain, mata yang lebih kecil untuk penglihatan periferal. Beberapa larva serangga, seperti ulat, memiliki berbagai jenis mata sederhana (stemmata) yang memberikan gambar kasar. Beberapa mata sederhana, yang disebut oselus, dapat ditemukan pada hewan seperti beberapa spesies siput, yang tidak benar-benar "melihat" dalam arti normal. Siput ini memiliki sel fotosensitif, tetapi tidak memiliki lensa dan tidak ada cara lain untuk memproyeksikan gambar ke sel-sel ini. Siput dapat membedakan antara terang dan gelap, tetapi tidak lebih dari itu. Hal ini memungkinkan siput untuk menjaganya dari sinar matahari langsung. Pada organisme yang hidup di dekat ventilasi hidrotermal, mata majemuk telah disederhanakan secara sekunder dan beradaptasi untuk menandai sinar inframerah yang dihasilkan oleh ventilasi panas, dengan cara ini mereka dapat menandai air panas dan menghindari dirinya terebus hidup-hidup.[11]
Jenis
Ada sepuluh jenis susunan mata—bahkan setiap metode teknologi untuk menangkap gambar optik yang biasa digunakan oleh manusia, dengan pengecualian lensa variabel dan lensa Fresnel, terdapat di alam.[3] Jenis-jenis mata dapat dikategorikan ke dalam "mata sederhana", dengan satu permukaan cekung fotoreseptif, dan "mata majemuk", yang terdiri dari sejumlah lensa individu diletakkan pada permukaan yang cembung.[3] Perhatikan bahwa "sederhana" tidak berarti penurunan tingkat kompleksitas atau ketajaman. Memang, setiap jenis mata dapat disesuaikan dengan hampir semua perilaku atau lingkungan. Satu-satunya keterbatasan khusus untuk jenis-jenis mata adalah resolusi—fisik dari mata majemuk mencegah untuk mencapai resolusi yang lebih baik dari 1°. Juga, mata superposisi dapat mencapai sensitivitas yang lebih besar dari mata aposisi, sehingga lebih cocok untuk makhluk yang hidup di tempat gelap.[3] Mata juga dikelompokkan ke dalam dua kelompok berdasarkan konstruksi seluler fotoreseptor, dengan sel fotoreseptor bersilia (seperti pada vertebrata) atau rabdomerik. Kedua kelompok ini tidak monofiletik; cnidaria juga memiliki sel bersilia,[12] dan beberapa annelida memiliki keduanya.[13]
Mata nonmajemuk
Keberadaan mata sederhana cukup tersebar luas dan bantalan lensa mata telah berevolusi setidaknya tujuh kali pada vertebrata, sefalopoda, annelida, krustasea, dan cubozoa.[14]
Mata ceruk

Mata ceruk, juga dikenal sebagai stemma, adalah bintik mata yang diatur ke dalam lubang untuk mengurangi sudut cahaya yang masuk dan memengaruhi bintik mata, memungkinkan organisme untuk menyimpulkan sudut cahaya yang masuk.[3] Ditemukan pada sekitar 85% dari filum hewan, bentuk-bentuk dasar yang mungkin adalah prekursor untuk jenis mata yang lebih maju dari "mata sederhana". Mata ceruk berukuran kecil, memiliki maksimal sekitar 100 sel yang berukuran sekitar 100 µm.[3] Direksionalitas dapat ditingkatkan dengan mengurangi ukuran bukaan, dengan menggabungkan lapisan reflektif di balik sel reseptor, atau dengan mengisi lubang dengan bahan refraktil.[3]
Mura telah mengembangkan lubang yang berfungsi sebagai mata dengan mengindra radiasi termal inframerah, di samping mata panjang gelombang optiknya seperti hewan vertebrata lainnya.
Mata berlensa sferis
Resolusi mata ceruk sangat bisa ditingkatkan dengan menggabungkan material yang memiliki indeks bias lebih tinggi untuk membentuk sebuah lensa, yang mungkin sangat mengurangi radius kabur yang dihadapi—karenanya meningkatkan resolusi yang diperoleh.[3] Bentuk yang paling dasar, terlihat pada beberapa spesies gastropoda dan annelida, terdiri dari sebuah lensa dengan satu indeks bias. Sebuah gambar yang jauh lebih tajam dapat diperoleh dengan menggunakan bahan dengan indeks bias tinggi yang nilainya menurun di sekitar tepi, bertujuan untuk mengurangi panjang fokus dan dengan demikian memungkinkan gambar yang tajam terbentuk pada retina.[3] Hal ini juga memungkinkan celah yang besar untuk memperoleh ketajaman gambar dengan memungkinkan lebih banyak cahaya yang masuk ke lensa, dan lensa datar dengan mengurangi aberasi sferis.[3] Seperti lensa heterogen yang diperlukan agar panjang fokus berkurang dari sekitar 4 kali radius lensa menjadi 2,5 radius lensa.[3]
Mata heterogen telah berevolusi setidaknya sembilan kali, empat kali atau lebih pada gastropoda, sekali pada copepoda, sekali pada annelida, sekali pada cumi,[3] dan sekali pada kiton, yang memiliki lensa aragonit.[15] Tidak ada organisme air yang masih memiliki lensa homogen, mungkin tekanan evolusioner untuk lensa heterogen cukup besar sehingga tahap ini menjadi cepat "teratasi".[3]
Mata ini menciptakan sebuah gambar yang cukup tajam, tetapi gerakan mata dapat menyebabkan kekaburan yang signifikan. Untuk meminimalkan pengaruh gerakan mata ketika hewan bergerak, sebagian besar mata telah menstabilkan otot matanya.[3]
Oselus serangga memiliki lensa sederhana, tetapi titik fokusnya selalu terletak di belakang retina; akibatnya mata ini tidak pernah bisa membentuk gambar yang tajam. Oselus (jenis mata ceruk pada artropoda) mengaburkan gambar di seluruh retina, dan karenanya sangat baik dalam menanggapi perubahan intensitas cahaya yang cepat di bidang pandang keseluruhan. Respons cepat ini lebih dipercepat lagi oleh berkas saraf yang besar, yang mengirimkan informasi ke otak.[16] Pemfokusan gambar juga akan menyebabkan citra matahari akan difokuskan pada beberapa reseptor, dengan kemungkinan kerusakan di bawah paparan cahaya yang kuat; melindungi reseptor akan menghalangi cahaya dan dengan demikian mengurangi sensitivitas.[16] Respons yang cepat ini telah membawa petunjuk bahwa oselus serangga digunakan terutama ketika terbang, karena oselus dapat digunakan untuk mendeteksi perubahan cahaya mendadak (karena cahaya, terutama sinar ultraviolet yang diserap oleh vegetasi, biasanya datang dari atas).[16]
Lensa ganda
Beberapa organisme laut memiliki lebih dari satu lensa, misalnya copepoda Pontella yang memiliki tiga lensa. Bagian luar memiliki permukaan parabola, melawan efek dari aberasi sferis yang memungkinkan pembentukan gambar tajam. Copepoda lain, Copilia, memiliki dua lensa di setiap mata, seperti yang diatur di dalam teleskop.[3] Pengaturan tersebut langka dan kurang dipahami, tetapi merupakan konstruksi alternatif. Beberapa lensa terlihat di beberapa hewan pemburu seperti elang dan laba-laba pelompat, memiliki kornea refraktif (dibahas berikutnya): struktur ini memiliki lensa negatif, memperbesar gambar yang diamati hingga 50% pada sel reseptor, sehingga meningkatkan resolusi optik.[3]
Kornea refraktif

Pada kebanyakan mata mamalia, burung, reptil, dan kebanyakan vertebrata darat lainnya (bersama dengan laba-laba dan beberapa larva serangga) cairan bening memiliki indeks bias lebih tinggi dari udara.[3] Secara umum, lensa tidak sferis. Lensa sferis menghasilkan aberasi sferis. Pada kornea refraktif, jaringan lensa dikoreksi dengan bahan lensa takhomogen (lihat lensa Luneburg), atau dengan bentuk taksferik.[3] Lensa datar memiliki kelemahan, yaitu kualitas penglihatan berkurang pada sumbu utama fokus. Dengan demikian, hewan yang telah berevolusi dengan bidang pandang luas sering memiliki mata yang menggunakan lensa takhomogen.[3]
Seperti disebutkan di atas, kornea refraktif hanya berguna di luar air. Di dalam air, terdapat sedikit perbedaan dalam indeks bias antara cairan bening dengan air di sekitarnya. Oleh karena itu, hewan yang telah kembali ke dalam air–misalnya penguin dan anjing laut–kehilangan kornea yang sangat melengkung dan kembali ke lensa berbasis penglihatan. Solusi alternatif yang dimiliki oleh beberapa hewan penyelam adalah memiliki kornea yang mampu memfokuskan sangat kuat.[3]
Mata pemantul

Sebuah alternatif untuk lensa adalah melapisi bagian dalam mata dengan "cermin", dan merefleksikan gambar agar memfokuskannya pada titik pusat.[3] Sifat mata ini memiliki arti bahwa jika seseorang mengintip ke pupil mata suatu organisme, seseorang itu akan melihat gambar yang sama dengan apa yang dilihat organisme karena gambar yang ditangkap dipantulkan kembali keluar.[3]
Banyak organisme kecil seperti rotifera, copepoda, dan cacing pipih menggunakan organ seperti ini, tetapi mata pemantul terlalu kecil untuk menghasilkan gambar yang dapat digunakan.[3] Beberapa organisme yang lebih besar, seperti kapis, juga menggunakan mata pemantul. Kapis Pecten hingga memiliki 100 mata pemantul per milimeter menyusuri tepi cangkangnya. Struktur ini dapat mendeteksi objek bergerak dengan melewati lensa berturutan.[3]
Terdapat setidaknya satu vertebrata, Dolichopteryx longipes, yang matanya termasuk optik reflektif untuk memfokuskan cahaya. Masing-masing dari dua mata pada seekor Dolichopteryx longipes mengumpulkan cahaya dari atas dan bawah. Cahaya yang datang dari atas difokuskan oleh lensa, sementara yang datang dari bawah difokuskan oleh cermin melengkung yang terdiri dari banyak lapisan lempeng kecil reflektif yang terbuat dari kristal guanin.[17]
Mata majemuk



Sebuah mata majemuk dapat terdiri dari ribuan fotoreseptor individual atau omatidia (omatidium, tunggal). Gambar yang dipersepsikan merupakan kombinasi ransangan dari berbagai omatidia ("satuan mata" individual) yang terletak di permukaan cembung, sehingga setiap omatidium menunjuk ke arah yang sedikit berbeda. Dibandingkan dengan mata sederhana, mata majemuk memiliki sudut pandang yang sangat besar, dapat mendeteksi gerakan cepat, dan dalam beberapa kasus dapat mendeteksi polarisasi cahaya.[18] Karena lensa individual sangat kecil, efek difraksi memaksakan batasan pada kemungkinan resolusi yang dapat diperoleh (dengan asumsi bahwa mata majemuk tidak berfungsi sebagai susunan berfase). Hal ini hanya dapat diatasi dengan meningkatkan ukuran dan jumlah lensa. Untuk melihat dengan resolusi sebanding dengan mata sederhana manusia, manusia akan membutuhkan mata majemuk sangat besar, memiliki radius sekitar 11 meter (36 kaki).[19]
Mata majemuk terbagi ke dalam dua kelompok: mata aposisi yang membentuk beberapa gambar terbalik, dan mata superposisi yang membentuk bayangan tunggal yang tegak.[20] Mata majemuk sering ditemukan pada artropoda, dan juga terdapat pada Annelida dan beberapa moluska dwikatup.[21] Mata majemuk, setidaknya pada artropoda, tumbuh dengan penambahan omatidia baru pada bagian tepi.[22]
Mata aposisi
Mata aposisi adalah bentuk mata yang paling umum, dan mungkin juga bentuk nenek moyang mata majemuk. Mata ini ditemukan di seluruh kelompok artropoda, meskipun mata ini mungkin telah berevolusi lebih dari sekali di dalam filum ini.[3] Beberapa annelida dan bivalvia juga memiliki mata aposisi. Mata ini juga dimiliki oleh Limulus, kepiting tapal kuda, dan terdapat petunjuk bahwa chelicerata lain mengembangkan mata sederhana dengan mereduksi titik awal mata majemuk[3] (beberapa ulat tampaknya telah mengembangkan mata majemuk dari mata sederhana dengan cara yang sebaliknya).
Mata aposisi bekerja dengan mengumpulkan sejumlah gambar, satu dari setiap mata, dan menggabungkannya di otak, dengan setiap mata biasanya memberi kontribusi satu titik informasi. Keunikan yang dimiliki mata aposisi adalah lensa yang memusatkan cahaya dari satu arah pada rabdom, sedangkan cahaya dari arah lain diserap oleh dinding gelap omatidium.
Mata superposisi
Mata superposisi terbagi menjadi tiga jenis, yaitu: mata superposisi pembiasan, pemantulan, dan parabolik. Mata superposisi pembiasan memiliki celah antara lensa dan rabdom, dan tidak memiliki dinding samping. Setiap lensa mengambil cahaya dengan sudut tertentu ke sumbu lensa dan memantulkannya dengan besar sudut yang sama di sisi lain. Hasilnya adalah sebuah gambar pada setengah radius mata, di situlah ujung rabdom berada. Jenis mata majemuk ini biasanya ditemukan pada serangga nokturnal karena dapat membuat gambar hingga 1000 kali lebih terang daripada gambar yang dibentu mata aposisi yang setara, meski dengan pengurangan resolusi.[23]
Dekapoda bertubuh panjang seperti udang, udang air tawar, lobster, dan lobster air tawar memiliki mata superposisi pemantulan, yang juga memiliki celah transparan namun menggunakan cermin bersudut dan bukan lensa.
Pada tipe mata majemuk superposisi parabolik, yang ditemukan pada artropoda seperti serangga pada ordo Ephemeroptera, permukaan bagian dalam berupa parabola dari setiap faset memfokuskan cahaya dari reflektor ke susunan sensor. Jenis mata ini berfungsi dengan membiaskan cahaya, lalu menggunakan cermin parabola untuk memfokuskan gambar, menggabungkan sifat mata aposisi dan superposisi.[11]
Lainnya
Jenis mata majemuk lainnya, yang ditemukan pada jantan ordo Strepsiptera, menggunakan serangkaian mata sederhana—mata dengan satu celah memberikan cahaya untuk retina pembentuk citra keseluruhan. Beberapa dari lubang ini bersama-sama membentuk mata majemuk strepsipteran, mirip dengan mata majemuk 'skizokroal' dari beberapa trilobita.[24] Karena masing-masing lubang mata adalah mata sederhana, mata ini akan menghasilkan gambar terbalik. Gambar tersebut digabungkan di otak untuk membentuk satu citra terpadu. Karena bukaan lubang lebih besar jika dibandingkan dengan mata majemuk, pengaturan ini memungkinkan penglihatan pada tingkat cahaya rendah.[3]

Penerbang baik seperti lalat atau lebah madu, atau serangga pemakan mangsa seperti belalang sentadu atau capung, memiliki omatidia dengan zona terspesialisasi disusun menjadi wilayah fovea yang memberikan penglihatan tajam. Pada wilayah penglihatan tajam ini, mata diratakan dan faset lebih besar. Perataan tersebut memungkinkan lebih banyak omatidia untuk menerima cahaya dari suatu tempat dan oleh karena itu memiliki resolusi yang lebih tinggi. Bintik hitam yang bisa dilihat pada mata majemuk beberapa serangga, selalu terlihat langsung, disebut pseudopupil. Pseudopupil ini terbentuk karena omatidia yang memiliki pengamatan terhadap sumbu optisnya menyerap sinar datang, sementara sisi lainnya memantulkan cahaya.[25]
Terdapat beberapa pengecualian dari jenis yang telah disebutkan di atas. Beberapa serangga memiliki mata yang disebut mata majemuk berlensa tunggal, sebuah tipe transisi antara mata majemuk multilensa tipe superposisi dengan mata berlensa tunggal yang ditemukan pada hewan dengan mata sederhana. Udang opossum Dioptromolis paucispinosa memiliki mata superposisi pembiasan, dengan hal yang jarang yaitu pada setiap mata terdapat faset tunggal yang besar dengan diameter tiga kali diameter faset lainnya dan di belakang faset ini terdapat kerucut kristalin yang besar. Struktur ini memproyeksikan citra tegak lurus pada retina terspesialisasi. Mata yang dihasilkan merupakan perpaduan mata sederhana di dalam mata majemuk.
Versi lain adalah mata pseudofaset, seperti yang terdapat pada Scutigera. Jenis mata ini terdiri dari gugus oselus di setiap sisi kepala, diatur sedemikian rupa sehingga menyerupai mata majemuk sejati.
Tubuh Ophiocoma wendtii, sejenis Ophiuroidea, ditutupi omatidia, mengubah seluruh kulitnya menjadi mata majemuk. Hal yang sama berlaku pada banyak kiton. Kaki tabung bulu babi mengandung protein fotoreseptor, yang bersama-sama bertindak sebagai mata majemuk. Struktur ini hanya memiliki sedikit pigmen penyaringan, tetapi dapat mendeteksi arah cahaya dari bayangan yang diberikan oleh tubuh legapnya.[26]
Evolusi
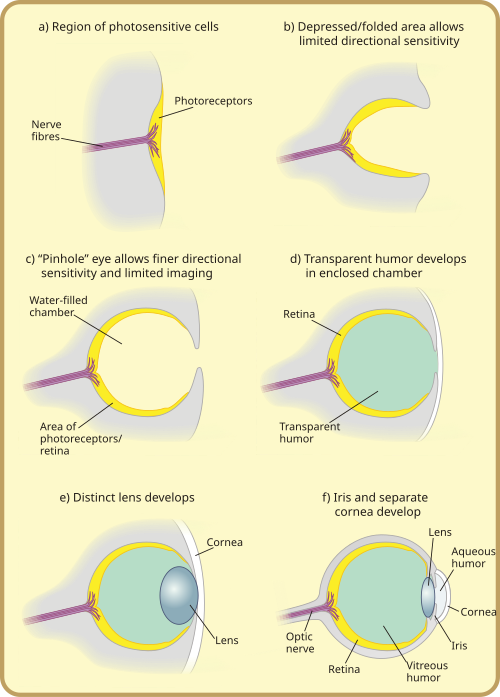
Fotoreseptor secara filogenetik sangat tua, dengan berbagai teori filogenesis.[27] Asal-usul bersama (monofili) semua mata hewan sekarang diterima secara luas sebagai fakta. Hal ini didasarkan pada ciri genetik bersama dari semua mata. Artinya, semua mata modern, bervariasi seperti semula, asal-usulnya berasal dari mata proto yang diyakini berevolusi sekitar 540 juta tahun yang lalu,[28][29][30] dan gen PAX6 dianggap sebagai faktor kunci dalam hal ini. Sebagian besar kemajuan pada mata proto diyakini telah memakan waktu beberapa juta tahun untuk berkembang, karena predator pertama yang mendapatkan pencitraan sejati akan menyentuh "perlombaan senjata"[31] di antara semua spesies yang tidak menjauh dari lingkungan fotopik. Hewan pemangsa dan predator yang bersaing sama-sama memiliki kelemahan yang berbeda tanpa suatu kemampuan dan cenderung tidak bertahan dan bereproduksi. Oleh karena itu, beberapa tipe mata dan subtipenya berkembang secara paralel (kecuali beberapa kelompok, seperti vertebrata, yang dipaksa masuk ke lingkungan fotopik pada tahap akhir).
Mata pada berbagai hewan menunjukkan adaptasi terhadap kebutuhannya. Misalnya, mata seekor burung pemangsa memiliki ketajaman penglihatan yang jauh lebih besar dibandingkan dengan mata manusia, dan dalam beberapa kasus dapat mendeteksi radiasi ultraviolet. Bentuk mata yang berbeda, misalnya, vertebrata dan moluska adalah contoh evolusi paralel, terlepas dari keturunan leluhur mereka yang jauh. Konvergensi fenotipik dari geometri sefalopoda dan sebagian besar mata vertebrata menciptakan kesan bahwa mata vertebrata berevolusi dari pencitraan mata sefalopoda, tetapi tak seperti itu, peran siliari dan kelas opsin rabdomerik pada kedua hewan terbalik,[32] dan memiliki lensa kristalin yang berbeda.[33]
"Mata" paling awal, disebut bintik mata, adalah kumpulan sederhana protein fotoreseptor pada hewan uniseluler. Pada makhluk hidup multiseluler, bintik mata multisel berevolusi, secara fisik mirip dengan kumpulan reseptor pengecap dan pembau. Mata ini hanya bisa mengindera kecerahan di sekitarnya, membedakan terang dan gelap, tetapi bukan arah sumber cahaya.[3]
Melalui perubahan bertahap, bintik mata pada spesies yang tinggal di lingkungan terang tertekan membentuk cekungan yang tidak dalam, sehingga memiliki sedikit kemampuan untuk membedakan kecerahan terarah yang diperoleh dengan menggunakan sudut cahaya yang menimpa sel tertentu untuk mengidentifikasi sumbernya. Lubang yang diperdalam dari waktu ke waktu, ukuran bukaan berkurang, dan jumlah sel fotoreseptor meningkat, membentuk kamera lubang jarum yang efektif dan mampu dengan samar-samar membedakan suatu bentuk.[34] Namun, nenek moyang remang modern, yang dianggap sebagai protovertebrata,[32] ternyata terdesak ke perairan dalam dan gelap, wilayah yang kurang rentan untuk terlihat oleh predator dan wilayah yang menguntungkan untuk memiliki bintik mata konveks, mengumpulkan lebih banyak cahaya dibandingkan dengan bintik mata datar ataupun cekung. Hal ini akan menyebabkan lintasan evolusioner yang agak berbeda pada mata vertebrata dibandingkan dengan mata hewan lainnya.
Pertumbuhan berlebih sel tipis dari sel transparan di atas bukaan mata, yang semula dibentuk untuk mencegah kerusakan pada bintik mata, memungkinkan cairan terpisah pada ruang mata terspesialisasi menjadi humor transparan yang mengoptimalkan penyaringan warna, menghambat radiasi berbahaya, memperbaiki indeks bias mata, dan memungkinkan fungsi di luar air. Sel pelindung transparan ini akhirnya terbagi menjadi dua lapisan, dengan cairan sirkulasi di antaranya memungkinkan sudut pandang yang lebih luas dan resolusi pencitraan yang lebih besar, dan ketebalan lapisan transparan meningkat secara bertahap, pada kebanyakan spesies dengan protein kristalin transparan.[35]
Celah antara lapisan jaringan secara alami membentuk bikonveks, struktur ideal secara optimal untuk memperoleh indeks bias yang normal. Secara independen, lapisan transparan dan lapisan nontransparan terpisah dari lensa membentuk kornea dan iris. Kemudian, pemisahan lapisan kembali terjadi membentuk humor, beranda depan. Humor ini mampu meningkatkan daya refraktif dan mengurangi masalah sirkulasi darah. Pembentukan cincin nontransparan memungkinkan lebih banyak pembuluh darah, lebih banyak sirkulasi, dan ukuran mata yang lebih besar.[35]
Hubungan dengan kebutuhan hidup
Mata umumnya beradaptasi dengan kebutuhan hidup organisme dan lingkungan yang menyandangnya. Misalnya, distribusi fotoreseptor cenderung sesuai dengan area yang membutuhkan ketajaman tinggi, dengan organisme pemindai horison, seperti makhluk hidup di dataran Afrika, memiliki garis horisontal ganglia dengan kepadatan tinggi, sedangkan makhluk hidup yang bertempat tinggal di pohon yang membutuhkan penglihatan serba baik cenderung memiliki distribusi ganglia simetris, dengan ketajaman menurun saat semakin jauh dari pusat.
Tentu saja, untuk kebanyakan jenis mata, tidak mungkin menyimpang dari bentuk bola, sehingga hanya densitas reseptor optik yang bisa diubah. Pada organisme dengan mata majemuk, jumlah omatidia dan bukan ganglia yang mencerminkan daerah dengan perolehan cahaya tertinggi.[3]: 23–4 Mata superposisi dibatasi pada bentuk bola, tetapi mata majemuk jenis lainnya dapat berubah menjadi bentuk yang memiliki lebih banyak omatidia yang selaras dengan, katakanlah, cakrawala, tanpa mengubah ukuran atau kepadatan omatidia individu.[36] Mata organisme pemindai horison memiliki tangkai sehingga bisa dengan mudah diselaraskan dengan cakrawala ketika berada di tempat miring, misalnya jika hewan itu berada di lereng.[25]

Perpanjangan dari konsep ini adalah bahwa mata predator biasanya memiliki zona penglihatan yang sangat tajam pada pusatnya, untuk membantu identifikasi mangsa.[36] Pada organisme perairan dalam, mungkin bukan pusat mata yang membesar. Amphipoda hyperiidea adalah hewan perairan dalam yang memakan organisme di atas mereka. Mata mereka hampir terbagi menjadi dua, dengan daerah bagian atas diperkirakan terlibat dalam mendeteksi siluet mangsa potensial—atau pemangsa—terhadap cahaya langit yang samar di atasnya. Dengan demikian, hyperiidea pada perairan yang lebih dalam, dengan perbandingan antara cahaya dan bayangan siluet sulit dibedakan, memiliki "mata bagian atas" yang lebih besar, dan mungkin kehilangan mata bagian bawah.[36] Persepsi kedalaman dapat ditingkatkan dengan memiliki mata yang membesar pada satu arah, sedikit mendistorsi mata memungkinkan jarak objek dapat diperkirakan dengan tingkat akurasi yang tinggi.[11]
Ketajaman lebih tinggi dimiliki oleh organisme jantan yang kawin di udara, karena organisme jantan ini harus bisa mengenali dan menilai calon pasangan dengan latar yang sangat besar.[36] Di sisi lain, mata organisme yang dipakai pada tingkat cahaya rendah, seperti sekitar fajar dan senja atau di perairan dalam, cenderung lebih besar untuk meningkatkan jumlah cahaya yang bisa ditangkap.[36]
Bukan hanya bentuk mata yang mungkin berdampak pada gaya hidup. Mata bisa menjadi bagian organisme yang paling terlihat, dan ini bisa bertindak sebagai tekanan pada organisme yang memiliki mata yang lebih transparan dengan biaya fungsi.[36]
Mata dapat terpasang pada tangkai untuk memberikan penglihatan serba lebih baik, dengan mengangkatnya ke atas karapas organisme. Hal ini juga memungkinkan mereka untuk melacak predator atau mangsa tanpa menggerakkan kepala.[11]
Fisiologi
Ketajaman penglihatan

Ketajaman penglihatan, atau daya pisah, adalah "kemampuan untuk membedakan detail halus" dan merupakan sifat dari sel kerucut.[37] Ketajaman penglihatan sering diukur dalam siklus per derajat, mengukur resolusi sudut, atau seberapa jauh mata dapat membedakan satu objek dengan objek lain dari segi sudut penglihatan. Resolusi dalam siklus per derajat dapat diukur menggunakan grafik batang dengan perbedaan jumlah siklus garis putih/hitam. Misalnya, jika masing-masing pola memiliki lebar 1,75 cm dan ditempatkan pada jarak 1 m dari mata, akan membentuk sudut 1 derajat, sehingga jumlah pasangan garis putih/hitam pada pola akan menjadi ukuran siklus per derajat pola itu. Angka tertinggi yang bisa dilihat oleh mata sebagai garis-garis terpisah, atau yang membedakan dari blok abu-abu, adalah pengukuran ketajaman penglihatan.
Untuk mata manusia dengan ketajaman yang sangat baik, resolusi teoretis maksimum adalah 50 siklus per derajat[38] (1,2 menit busur per pasangan garis, atau 0,35 mm pasangan garis pada jarak 1 m). Seekor tikus hanya mampu memisahkan sekitar 1 sampai 2 siklus per derajat.[39] Seekor kuda memiliki ketajaman yang lebih tinggi melalui sebagian besar bidang penglihatan matanya dibandingkan dengan yang dimiliki manusia, tetapi tidak serupa dengan ketajaman daerah fovea pada pusat mata manusia.[40]
Aberasi sferis membatasi resolusi pupil berdiameter 7 mm hingga sekitar 3 menit busur per pasangan garis. Pada pupil berdiameter 3 mm, aberasi sferis sangat berkurang, meningkatkan resolusi sekitar 1,7 menit busur per pasangan garis.[41] Resolusi 2 menit busur per pasangan garis, setara dengan celah 1 menit busur pada optotipe, sesuai dengan 20/20 (penglihatan normal) pada manusia.
Namun, resolusi pada mata majemuk berkaitan dengan ukuran omatidia tunggal dan jarak antar omatidia yang bersebelahan. Secara fisik ukuran omatidia tidak dapat dikurangi untuk mencapai ketajaman seperti yang terlihat dengan mata berlensa tunggal pada mamalia. Mata majemuk memiliki ketajaman yang jauh lebih rendah dibandingkan dengan mata vertebrata.[42]
Persepsi warna
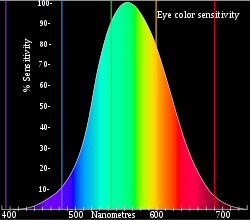
Penglihatan warna adalah kemampuan organisme untuk membedakan cahaya dengan kualitas spektral yang berbeda.[43] Semua organisme terbatas pada rentang spektrum elektromagnetik yang sempit, bervariasi antar makhluk hidup, tetapi sebagian besar mampu melihat panjang gelombang antara 400 hingga 700 nm.[44] Rentang ini merupakan bagian yang kecil dari spektrum elektromagnetik, mungkin mencerminkan evolusi organ dari makhluk hidup bawah laut: air menghalangi semua kecuali dua jendela kecil spektrum elektromagnetik, dan tidak ada tekanan evolusioner pada hewan darat untuk memperluas rentang ini.[45]
Pigmen yang paling sensitif, rodopsin, memiliki respons puncak pada 500 nm.[46] Perubahan kecil pada gen yang mengkodekan protein ini dapat mengubah respons puncak beberapa nm.[4] Pigmen pada lensa juga dapat menyaring cahaya masuk dan mengubah respons puncak.[4] Banyak organisme tidak dapat membedakan warna, melihat dengan warna abu-abu, yang menunjukkan bahwa penglihatan warna memerlukan berbagai sel pigmen yang terutama sensitif terhadap rentang spektrum yang lebih kecil. Pada primata, tokek, dan organisme lainnya, sel pigmen ini berbentuk sel kerucut, yang kemudian sel batang yang lebih sensitif berevolusi dari sel ini.[46] Bahkan, jika organisme secara fisik mampu membedakan warna yang berbeda, hal ini tidak berarti bahwa organisme tersebut dapat merasakan warna yang berbeda, hanya dengan tes perilaku hal ini dapat disimpulkan.[4]
Kebanyakan organisme dengan penglihatan warna mampu mendeteksi sinar ultraviolet. Cahaya dengan energi tinggi ini mampu merusak sel reseptor. Dengan beberapa pengecualian (ular dan mamalia berplasenta), kebanyakan organisme menghindari efek ini dengan memiliki tetesan minyak penyerap di sekitar sel kerucutnya. Alternatifnya, organisme yang telah kehilangan tetesan minyak ini dalam perjalanan evolusinya membuat lensa yang tahan terhadap sinar ultraviolet, menghalangi kemungkinan adanya sinar ultraviolet yang terdeteksi hingga tidak sampai ke retina.[46]
Sel batang dan kerucut
Retina memiliki dua jenis sel fotoreseptor mayor yang peka cahaya dan digunakan untuk penglihatan: sel batang dan sel kerucut. Sel batang tidak dapat membedakan warna, tetapi bertanggung jawab dalam penglihatan pada cahaya rendah (skotopik) monokrom (hitam putih). Sel batang bekerja dengan baik pada cahaya redup karena mengandung pigmen, rodopsin (ungu), yang sensitif pada intensitas cahaya rendah, tetapi jenuh pada intensitas yang lebih tinggi (fotopik). Sel batang tersebar ke seluruh retina kecuali fovea dan bintik buta. Kepadatan sel batang lebih besar pada retina perifer dibandingkan pada retina sentral.
Sel kerucut bertanggung jawab dalam penglihatan warna. Sel ini membutuhkan cahaya yang lebih terang untuk berfungsi dibandingkan dengan cahaya yang dibutuhkan sel batang. Pada manusia, terdapat tiga jenis sel kerucut, yaitu: sel kerucut yang sangat peka terhadap panjang gelombang panjang, panjang gelombang menengah, dan panjang gelombang pendek (sering disebut sebagai merah, hijau, dan biru secara berurutan, meskipun puncak sensitivitasnya tidak benar-benar pada warna ini). Warna yang terlihat adalah gabungan efek stimulus dan respons dari ketiga jenis sel kerucut ini. Sel kerucut sebagian besar terkonsentrasi di dan dekat fovea. Hanya sedikit yang terdapat di sisi retina. Objek terlihat paling tajam pada fokus ketika gambar jatuh di fovea, seperti ketika seseorang melihat objek secara langsung. Sel kerucut dan batang dihubungkan melalui sel antara di retina ke serabut saraf optik. Ketika sel batang dan kerucut dirangsang oleh cahaya, keduanya terhubung melalui sel yang berdampingan di dalam retina untuk mengirim sinyal listrik ke serabut saraf optik. Saraf optik mengirimkan impuls melalui serabut saraf ini ke otak.[46]
Pigmentasi
Molekul pigmen yang digunakan mata bervariasi, tetapi perbedaan ini dapat digunakan untuk menentukan jarak evolusioner antara kelompok yang berbeda, dan juga dapat menjadi bantuan dalam menentukan kelompok mana yang terkait erat–walaupun terdapat masalah konvergensi.[46]
Opsin adalah pigmen yang terlibat dalam fotoresepsi. Pigmen lain, seperti melanin, digunakan untuk melindungi sel fotoreseptor dari cahaya yang lolos dari samping. Kelompok protein opsin berevolusi jauh sebelum nenek moyang terakhir hewan, dan terus melakukan diversifikasi sejak saat itu.[4]
Terdapat dua jenis opsin yang terlibat dalam penglihatan, yaitu c-opsin yang berasosiasi dengan sel fotoreseptor bersilia dan r-opsin yang berasosiasi dengan sel fotoreseptor rabdomer.[47] Mata vertebrata biasanya mengandung sel bersilia dengan c-opsin dan mata invertebrata (bilaterian) memiliki sel rabdomer dengan r-opsin. Namun, beberapa sel ganglion vertebrata mengekspresikan r-opsin, menunjukkan bahwa nenek moyang vertebrata menggunakan pigmen ini dalam penglihatan, dan sisa-sisa itu bertahan di mata.[47] Demikian juga, telah ditemukan bahwa c-opsin diekspresikan di otak beberapa invertebrata. C-opsin ini mungkin telah diekspresikan oleh sel bersilia pada mata larva, yang kemudian diserap ke otak pada metamorfosis ke bentuk dewasa.[47] C-opsin juga ditemukan di beberapa mata invertebrata bilaterian yang diturunkan, seperti mata palium dari moluska bivalvia, tetapi mata lateral (yang mungkin merupakan tipe nenek moyang untuk kelompok ini, jika mata berevolusi sekali) selalu menggunakan r-opsin.[47] Cnidaria, yang merupakan kelompok luar dari taksa yang disebutkan di atas, mengekspresikan c-opsin–namun r-opsin belum ditemukan di dalam kelompok ini.[47] Kebetulan, melanin pada cnidaria diproduksi dengan cara yang sama seperti pada vertebrata, menunjukkan penurunan pigmen ini.[47]
Lihat pula
- Adaptasi (mata)
- Mata artropoda
- Mata mamalia
- Mata manusia
- Mata moluska
- Mata sederhana (invertebrata)
- Mata sefalopoda
- Perkembangan mata
Catatan dan referensi
- ^ Tidak ada konsensus universal pada jumlah yang tepat dari filum Animalia; penyataan jumlah sedikit bervariasi dari penulis ke penulis.
- ^ Arti kata GALAT! URL tidak ditemukan atau tidak sah. dalam situs web {{{ver}}} oleh lembaga penyusun kamus.
- ^ Arti kata GALAT! URL tidak ditemukan atau tidak sah. dalam situs web {{{ver}}} oleh lembaga penyusun kamus.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad Land, M. F.; Fernald, R. D. (1992). "The evolution of eyes". Annual Review of Neuroscience. 15: 1–29. doi:10.1146/annurev.ne.15.030192.000245. PMID 1575438.
- ^ a b c d e Frentiu, Francesca D.; Adriana D. Briscoe (2008). "A butterfly eye's view of birds". BioEssays. 30 (11–12): 1151–62. doi:10.1002/bies.20828. PMID 18937365.
- ^ "Circadian Rhythms Fact Sheet". National Institue of General Medical Sciences. Diarsipkan dari asli tanggal 2020-03-13. Diakses tanggal 3 June 2015.
- ^ Breitmeyer, Bruno (2010). Blindspots: The Many Ways We Cannot See. New York: Oxford University Press. hlm. 4. ISBN 978-0-19-539426-9.
- ^ Nairne, James (2005). Psychology. Belmont: Wadsworth Publishing. ISBN 0-495-03150-X. OCLC 61361417.
- ^ Bruce, Vicki; Green, Patrick R.; Georgeson, Mark A. (1996). Visual Perception: Physiology, Psychology and Ecology. Psychology Press. hlm. 20. ISBN 0-86377-450-4.
- ^ BioMedia Associates Educational Biology Site: What animal has a more sophisticated eye, Octopus or Insect? Diarsipkan 2008-03-05 di Wayback Machine.
- ^ "Who You Callin' "Shrimp"? – National Wildlife Magazine". Nwf.org. 2010-10-01. Diarsipkan dari asli tanggal 2010-08-09. Diakses tanggal 2014-04-03.
- ^ a b c d Cronin, T. W.; Porter, M. L. (2008). "Exceptional Variation on a Common Theme: the Evolution of Crustacean Compound Eyes". Evolution Education and Outreach. 1 (4): 463–475. doi:10.1007/s12052-008-0085-0.
- ^ Kozmik, Z.; Ruzickova, J.; Jonasova, K.; Matsumoto, Y.; Vopalensky, P.; Kozmikova, I.; Strnad, H.; Kawamura, S.; Piatigorsky, J.; Paces, V.; Vlcek, C. (Jul 2008). "Assembly of the cnidarian camera-type eye from vertebrate-like components" (Free full text). Proceedings of the National Academy of Sciences of the United States of America. 105 (26): 8989–8993. Bibcode:2008PNAS..105.8989K. doi:10.1073/pnas.0800388105. ISSN 0027-8424. PMC 2449352. PMID 18577593.
- ^ Fernald, Russell D. (2006). "Casting a Genetic Light on the Evolution of Eyes" (PDF). Science. 313 (5795): 1914–1918. Bibcode:2006Sci...313.1914F. doi:10.1126/science.1127889. PMID 17008522. Diarsipkan dari asli (PDF) tanggal 2016-03-05. Diakses tanggal 2017-02-07.
- ^ Nilsson, Dan-E. (1989). "Vision optics and evolution". BioScience. 39 (5): 298–307. doi:10.2307/1311112. JSTOR 1311112.
- ^ Speiser, D. I.; Eernisse, D. J.; Johnsen, S. N. (2011). "A Chiton Uses Aragonite Lenses to Form Images". Current Biology. 21 (8): 665–670. doi:10.1016/j.cub.2011.03.033. PMID 21497091.
- ^ a b c Wilson, M. (1978). "The functional organisation of locust ocelli". Journal of Comparative Physiology. 124 (4): 297–316. doi:10.1007/BF00661380.
- ^ Wagner, H.J.; Douglas, R.H.; Frank, T.M.; Roberts, N.W.; Partridge, J.C. (Jan 27, 2009). "A Novel Vertebrate Eye Using Both Refractive and Reflective Optics". Current Biology. 19 (2): 108–114. doi:10.1016/j.cub.2008.11.061. PMID 19110427.
- ^ Völkel, R; Eisner, M; Weible, K. J (June 2003). "Miniaturized imaging systems" (PDF). Microelectronic Engineering. 67–68 (1): 461–472. doi:10.1016/S0167-9317(03)00102-3. Diarsipkan dari asli (PDF) tanggal 2008-10-01. Diakses tanggal 2017-03-21.
- ^ Land, Michael (1997). "Visual Acuity in Insects" (PDF). Annual Review of Entomology. 42: 147–77. doi:10.1146/annurev.ento.42.1.147. PMID 15012311. Diarsipkan dari asli (PDF) tanggal 2004-11-23. Diakses tanggal 27 Mei 2013.
- ^ Gaten, Edward (1998). "Optics and phylogeny: is there an insight? The evolution of superposition eyes in the Decapoda (Crustacea)". Contributions to Zoology. 67 (4): 223–236. Diarsipkan dari asli tanggal 2013-01-12. Diakses tanggal 2017-03-21.
- ^ Ritchie, Alexander (1985). "Ainiktozoon loganense Scourfield, a protochordate? from the Silurian of Scotland". Alcheringa. 9 (2): 137. doi:10.1080/03115518508618961.
- ^ Mayer, G. (2006). "Structure and development of onychophoran eyes: What is the ancestral visual organ in arthropods?". Arthropod Structure and Development. 35 (4): 231–245. doi:10.1016/j.asd.2006.06.003. PMID 18089073.
- ^ Greiner, Birgit (16 Desember 2005). Adaptations for nocturnal vision in insect apposition eyes (PDF) (Ph.D.). Lund University. Diakses tanggal 13 November 2014. "Salinan arsip" (PDF). Diarsipkan dari asli (PDF) tanggal 2013-02-09. Diakses tanggal 2017-05-17.
- ^ Horváth, Gábor; Clarkson, Euan N.K. (1997). "Survey of modern counterparts of schizochroal trilobite eyes: Structural and functional similarities and differences". Historical Biology. 12.
- ^ a b Jochen Zeil; Maha M. Al-Mutairi (1996). "Variations in the optical properties of the compound eyes of Uca lactea annulipes" (PDF). The Journal of Experimental Biology. 199 (7): 1569–1577. PMID 9319471. Diarsipkan dari asli (PDF) tanggal 2009-02-25. Diakses tanggal 2017-05-21.
- ^ Ullrich-Luter, E. M.; Dupont, S.; Arboleda, E.; Hausen, H.; Arnone, M. I. (2011). "Unique system of photoreceptors in sea urchin tube feet". Proceedings of the National Academy of Sciences. 108 (20): 8367–8372. doi:10.1073/pnas.1018495108. PMC 3100952. PMID 21536888.
- ^ Autrum, H. "Introduction". Dalam H. Autrum (ed.). Comparative Physiology and Evolution of Vision in Invertebrates- A: Invertebrate Photoreceptors. Handbook of Sensory Physiology. Vol. VII/6A. New York: Springer-Verlag. hlm. 4, 8–9. ISBN 3-540-08837-7.
- ^ Halder, G.; Callaerts, P.; Gehring, W.J. (1995). "New perspectives on eye evolution". Curr. Opin. Genet. Dev. 5 (5): 602–609. doi:10.1016/0959-437X(95)80029-8. PMID 8664548.
- ^ Halder, G.; Callaerts, P.; Gehring, W.J. (1995). "Induction of ectopic eyes by targeted expression of the eyeless gene in Drosophila". Science. 267 (5205): 1788–1792. Bibcode:1995Sci...267.1788H. doi:10.1126/science.7892602. PMID 7892602.
- ^ Tomarev, S.I.; Callaerts, P.; Kos, L.; Zinovieva, R.; Halder, G.; Gehring, W.; Piatigorsky, J. (1997). "Squid Pax-6 and eye development". Proc. Natl. Acad. Sci. USA. 94 (6): 2421–2426. Bibcode:1997PNAS...94.2421T. doi:10.1073/pnas.94.6.2421. PMC 20103. PMID 9122210.
- ^ Conway-Morris, S. (1998). The Crucible of Creation. Oxford: Oxford University Press
- ^ a b Trevor D. Lamb; Shaun P. Collin; Edward N. Pugh Jr. (2007). "Evolution of the vertebrate eye: opsins, photoreceptors, retina and eye cup". Nature Reviews Neuroscience. 8 (12): 960–976. doi:10.1038/nrn2283. PMC 3143066. PMID 18026166.
- ^ Staaislav I. Tomarev; Rina D. Zinovieva (1988). "Squid major lens polypeptides are homologous to glutathione S-transferases subunits". Nature. 336 (6194): 86–88. Bibcode:1988Natur.336...86T. doi:10.1038/336086a0. PMID 3185725.
- ^ "Eye-Evolution?". Library.thinkquest.org. Diarsipkan dari asli tanggal 2012-09-15. Diakses tanggal 1 September 2012.
- ^ a b Fernald, Russell D. (2001). The Evolution of Eyes: Where Do Lenses Come From? Diarsipkan 2006-03-19 di Wayback Machine. Karger Gazette 64: "The Eye in Focus".
- ^ a b c d e f Land, M. F. (1989). "The eyes of hyperiid amphipods: relations of optical structure to depth". Journal of Comparative Physiology A. 164 (6): 751–762. doi:10.1007/BF00616747.
- ^ Ali & Klyne 1985, hlm. 28
- ^ Russ, John C. (2006). The Image Processing Handbook. CRC Press. ISBN 0-8493-7254-2. OCLC 156223054.
The upper limit (finest detail) visible with the human eye is about 50 cycles per degree,... (Fifth Edition, 2007, Page 94)
- ^ Klaassen, Curtis D. (2001). Casarett and Doull's Toxicology: The Basic Science of Poisons. McGraw-Hill Professional. ISBN 0-07-134721-6. OCLC 47965382.
- ^ C.R., Nave (2016). "The Retina". HyperPhysics. Department of Physics and Astronomy Georgia State University. Diarsipkan dari asli tanggal 2015-05-04. Diakses tanggal 20 Juni 2017.
- ^ Fischer, Robert E.; Tadic-Galeb, Biljana; Plympton, Rick (2000). Steve Chapman (ed.). Optical System Design. McGraw-Hill Professional. ISBN 0-07-134916-2. OCLC 247851267.
- ^ Barlow, H. B. (1952). "The size of ommatidia in apposition eyes". J Exp Biol. 29 (4): 667–674. Diarsipkan dari asli tanggal 2016-08-31. Diakses tanggal 2017-06-20.
- ^ Ali & Klyne 1985, hlm. 161
- ^ Barlow, Horace Basil; Mollon, J. D (1982). The Senses. Cambridge: Cambridge University Press. hlm. 98. ISBN 0-521-24474-9.
- ^ Fernald, Russell D. (1997). "The Evolution of Eyes". Brain, Behaviour and Evolution. 50 (4): 253–259. doi:10.1159/000113339. PMID 9310200. Diarsipkan dari asli (PDF) tanggal 2012-11-20. Diakses tanggal 2017-06-21.
- ^ a b c d e Goldsmith, T. H. (1990). "Optimization, Constraint, and History in the Evolution of Eyes". The Quarterly Review of Biology. 65 (3): 281–322. doi:10.1086/416840. JSTOR 2832368. PMID 2146698.
- ^ a b c d e f Nilsson, E.; Arendt, D. (Dec 2008). "Eye Evolution: the Blurry Beginning". Current Biology. 18 (23): R1096 – R1098. doi:10.1016/j.cub.2008.10.025. ISSN 0960-9822. PMID 19081043.
Bacaan tambahan
- Yong, Ed (14 January 2016). "Inside the Eye: Nature's Most Exquisite Creation". National Geographic.
Bibliografi
- Ali, Mohamed Ather; Klyne, M. A. (1985). Vision in Vertebrates. New York: Plenum Press. ISBN 0-306-42065-1.
Pranala luar
- Evolution of the eye
- Anatomy of the eye – flash animated interactive.
- Webvision. The organisation of the retina and visual system. An in-depth treatment of retinal function, open to all but geared most toward graduate students.
- Eye strips images of all but bare essentials before sending visual information to brain, UC Berkeley research shows
| Internasional | |
|---|---|
| Nasional | |
| Lain-lain | |
Konten ini disalin dari wikipedia, mohon digunakan dengan bijak.

