Asam amino nonproteinogenik

Dalam biokimia, asam amino non-kode atau asam amino non-proteinogenik berbeda dari 22 asam amino proteinogenik (21 pada eukariota[catatan 1]), yang secara alami dikodekan dalam genom organisme untuk perakitan protein. Namun, lebih dari 140 asam amino non-proteinogenik terjadi secara alami dalam protein (tetapi tidak termasuk dalam kode genetik) dan ribuan lainnya dapat terjadi di alam atau disintesis di laboratorium.[1] Asam amino yang disintesis secara kimia dapat disebut asam amino tidak alami. Asam amino tidak alami dapat disiapkan secara sintetis dari analog aslinya melalui modifikasi seperti alkilasi amina, substitusi rantai samping, siklisasi perpanjangan ikatan struktural, dan penggantian isosterik dalam tulang punggung asam amino.[2] Banyak asam amino non-proteinogenik yang penting:
- intermediet dalam biosintesis,
- dalam pembentukan protein pasca-translasi,
- berperan fisiologis (misalnya komponen dinding sel bakteri, neurotransmiter, dan toksin),
- senyawa farmakologis alami atau buatan manusia,
- terdapat dalam meteorit atau digunakan dalam eksperimen prebiotik (seperti percobaan Miller–Urey),
- mungkin merupakan neurotransmiter penting, seperti asam aminobutirat gamma,[3] dan
- dapat memainkan peran krusial dalam bioenergetika seluler, seperti kreatina.[4]
Definisi dengan negasi

Kode genetik mengkodekan 20 asam amino standar untuk dimasukkan ke dalam protein selama [[translasi (genetik)|translasi. Namun, terdapat dua asam amino proteinogenik tambahan: selenosistein dan pirolisin. Asam amino non-standar ini tidak memiliki kodon khusus, tetapi ditambahkan sebagai pengganti kodon stop ketika urutan spesifik hadir, kodon UGA dan elemen SECIS untuk selenosistein,[5] urutan hilir UAG PYLIS untuk pirolisin.[6] Semua asam amino lainnya disebut "non-proteinogenik".
-
 Selenosistein: Asam amino ini mengandung gugus selenol pada karbon β-nya.
Selenosistein: Asam amino ini mengandung gugus selenol pada karbon β-nya. -
 Pirolisin: Asam amino ini terbentuk dengan menggabungkan gugus ε-amino lisin dengan cincin pirolina yang dikarboksilasi.
Pirolisin: Asam amino ini terbentuk dengan menggabungkan gugus ε-amino lisin dengan cincin pirolina yang dikarboksilasi.

Terdapat berbagai kelompok asam amino:[7]
- 20 asam amino standar
- 22 asam amino proteinogenik
- lebih dari 80 asam amino yang dihasilkan secara abiotik dalam konsentrasi tinggi
- sekitar 900 diproduksi melalui jalur alami
- lebih dari 118 asam amino hasil rekayasa telah dimasukkan ke dalam protein
Kelompok-kelompok ini tumpang tindih, tetapi tidak identik. Semua 22 asam amino proteinogenik disintesis secara biologis oleh organisme dan beberapa (tetapi tidak semuanya) juga bersifat abiotik (ditemukan dalam eksperimen prebiotik dan meteorit). Beberapa asam amino alami seperti norleusin salah dimasukkan secara translasi ke dalam protein karena ketidakakuratan proses sintesis protein. Banyak asam amino seperti ornitina adalah perantara metabolisme yang diproduksi secara biosintetik, tetapi tidak dimasukkan secara translasi ke dalam protein. Modifikasi pascatranslasi residu asam amino dalam protein menyebabkan terbentuknya banyak asam amino proteinaceous, namun non-proteinogenik. Asam amino lainnya hanya ditemukan dalam campuran abiotik (misalnya α-metilnorvalina). Lebih dari 30 asam amino tidak alami telah dimasukkan secara translasi ke dalam protein dalam sistem rekayasa, namun tidak bersifat biosintetik.[7]
Tata nama
Selain sistem penomoran IUPAC untuk membedakan berbagai karbon dalam molekul organik, dengan secara berurutan memberikan nomor pada setiap karbon, termasuk yang membentuk gugus karboksil, karbon di sepanjang rantai samping asam amino juga dapat diberi label dengan huruf Yunani, di mana karbon α adalah karbon kiral pusat yang memiliki gugus karboksil, rantai samping, dan gugus amino pada asam α-amino — karbon dalam gugus karboksil tidak dihitung.[8]
Non-alfa

Beberapa asam amino non-α terdapat dalam organisme. Dalam struktur ini, gugus amina terletak lebih jauh dari ujung asam karboksilat molekul asam amino. Dengan demikian, asam amino β memiliki gugus amina yang terikat pada karbon kedua, dan asam amino γ memiliki gugus amina pada karbon ketiga. Contohnya termasuk β-alanin, GABA, dan asam δ-aminolevulinat.[butuh rujukan]
-
![β-alanina: asam amino yang dihasilkan oleh aspartat 1-dekarboksilase dan prekursor koenzim A[9] dan peptida karnosin dan anserin.](//upload.wikimedia.org/wikipedia/commons/thumb/3/35/Beta-alanine_structure.svg/120px-Beta-alanine_structure.svg.png)
-
 Asam aminobutirat gamma (GABA): neurotransmiter pada manusia dan hewan.
Asam aminobutirat gamma (GABA): neurotransmiter pada manusia dan hewan. -

-
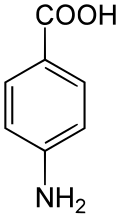 Asam 4-aminobenzoat (PABA): perantara dalam biosintesis asam folat
Asam 4-aminobenzoat (PABA): perantara dalam biosintesis asam folat
![β-alanina: asam amino yang dihasilkan oleh aspartat 1-dekarboksilase dan prekursor koenzim A[9] dan peptida karnosin dan anserin.](http://upload.wikimedia.org/wikipedia/commons/thumb/3/35/Beta-alanine_structure.svg/120px-Beta-alanine_structure.svg.png)


Alasan mengapa asam α-amino digunakan dalam protein telah dikaitkan dengan frekuensinya dalam meteorit dan eksperimen prebiotik.[10] Spekulasi awal mengenai sifat-sifat merugikan asam β-amino dalam hal struktur sekunder[10] ternyata tidak benar.[11]
Asam D-amino
Beberapa asam amino mengandung kiralitas absolut yang berlawanan, bahan kimia yang tidak tersedia dari mesin translasi dan transkripsi ribosom normal. Sebagian besar dinding sel bakteri dibentuk oleh peptidoglikan, polimer yang terdiri dari gula amino yang dihubungkan silang dengan oligopeptida pendek yang menjembatani satu sama lain. Oligopeptida disintesis secara non-ribosom dan mengandung beberapa kekhasan termasuk asam D-amino, umumnya D-alanina dan D-glutamat. Kekhasan lainnya adalah bahwa yang pertama dirasemisasi oleh enzim pengikat PLP (dikodekan oleh alr atau homolog dadX), sedangkan yang terakhir dirasemisasi oleh enzim independen kofaktor (murI). Beberapa varian hadir, pada bakteri Thermoga spp. D-Lisin hadir dan pada bakteri resisten vankomisin tertentu terdapat D-serin (gen vanT).[12][13]
Tanpa hidrogen pada karbon α
Semua asam amino proteinogenik memiliki setidaknya satu hidrogen pada karbon α. Glisin memiliki dua hidrogen, dan semua asam amino lainnya memiliki satu hidrogen dan satu rantai samping. Penggantian hidrogen yang tersisa dengan substituen yang lebih besar, seperti gugus metil, akan mendistorsi "tulang punggung" protein.[10]
Pada beberapa fungi, asam α-aminoisobutirat diproduksi sebagai prekursor peptida, beberapa di antaranya menunjukkan sifat antibiotik.[14]
-
 alanina
alanina -
 asam aminoisobutirat
asam aminoisobutirat -
 dehidroalanina
dehidroalanina



Stereosenter asam amino kembar
Sebagian dari asam amino L-α ambigu mengenai mana dari dua ujung yang merupakan karbon α. Dalam protein, residu sistein dapat membentuk ikatan disulfida dengan residu sistein lain, sehingga menghubungkan silang protein. Dua sistein yang terhubung silang membentuk molekul sistina. Sistein dan metionina umumnya diproduksi melalui sulfurilasi langsung, tetapi pada beberapa organisme, keduanya dapat diproduksi melalui transsulfurasi, di mana homoserina atau serina yang diaktifkan digabungkan dengan sistein atau homosistein membentuk sistationina. Senyawa serupa adalah lantionina, yang dapat dilihat sebagai dua molekul alanina yang bergabung melalui ikatan tioeter dan ditemukan di berbagai organisme. Demikian pula, asam jengkolat, toksin tumbuhan dari jengkol, terdiri dari dua sistein yang dihubungkan oleh gugus metilena. Asam diaminopimelat digunakan sebagai jembatan dalam peptidoglikan dan juga digunakan sebagai prekursor lisin (melalui dekarboksilasinya). [butuh rujukan]
-
 sistina
sistina -
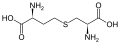 sistationina
sistationina -
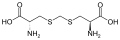
 lantionina
lantionina -
-
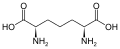 asam diaminopimelat
asam diaminopimelat
Asam amino prebiotik dan biokimia alternatif
Dalam meteorit dan dalam percobaan prebiotik (misalnya, percobaan Miller–Urey) ditemukan lebih banyak asam amino daripada dua puluh asam amino standar, beberapa di antaranya memiliki konsentrasi lebih tinggi daripada yang standar. Telah diperkirakan bahwa jika kehidupan berbasis asam amino muncul di tempat lain di alam semesta, tidak lebih dari 75% asam amino akan sama.[10]
| Molekul | Pelepasan listrik | Meteorit Murchison |
|---|---|---|
| glisina | 100 | 100 |
| alanina | 180 | 36 |
| asam α-amino-n-butirat | 61 | 19 |
| norvalina | 14 | 14 |
| valina | 4,4 | |
| norleusin | 1,4 | |
| leusin | 2,6 | |
| isoleusin | 1,1 | |
| aloisoleusin | 1,2 | |
| t-leusin | < 0,005 | |
| asam α-amino-n-heptanoat | 0,3 | |
| prolin | 0,3 | 22 |
| asam pipekolat | 0,01 | 11 |
| asam α,β-diaminopropionat | 1,5 | |
| asam α,γ-diaminobutirat | 7,6 | |
| ornitin | < 0,01 | |
| lisin | < 0,01 | |
| asam aspartat | 7,7 | 13 |
| asam glutamat | 1,7 | 20 |
| serina | 1,1 | |
| treonina | 0,2 | |
| alotreonina | 0,2 | |
| metionina | 0,1 | |
| homosistein | 0,5 | |
| homoserina | 0,5 | |
| β-alanina | 4,3 | 10 |
| asam β-amino-n-butirat | 0,1 | 5 |
| asam β-aminoisobutirat | 0,5 | 7 |
| asam aminobutirat gamma | 0,5 | 7 |
| asam α-aminoisobutirat | 7 | 33 |
| isovalina | 1 | 11 |
| sarkosina | 12,5 | 7 |
| N-etilglisina | 6,8 | 6 |
| N-propilglisina | 0,5 | |
| N-isopropilglisina | 0,5 | |
| N-metilalanina | 3,4 | 3 |
| N-etilalanina | < 0,05 | |
| N-metil-β-alanina | 1 | |
| N-etil-β-alanina | < 0,05 | |
| isoserina | 1,2 | |
| asam α-hidroksi-γ-aminobutirat | 17 |
Rantai samping lurus
Kode genetik telah digambarkan sebagai "kecelakaan yang membeku" dan alasan mengapa hanya ada satu asam amino standar dengan rantai lurus, alanina, mungkin hanya redundansi dengan valina, leusin, dan isoleusin.[10] Namun, asam amino rantai lurus dilaporkan membentuk heliks alfa yang jauh lebih stabil.[15]
-
 glisina (rantai samping hidrogen)
glisina (rantai samping hidrogen) -
alanina (rantai samping metil)
-
 homoalanina, atau asam α-aminobutirat (rantai samping etil)
homoalanina, atau asam α-aminobutirat (rantai samping etil) -
 norvalina (rantai samping n-propil)
norvalina (rantai samping n-propil) -
 norleusina (rantai samping n-butil)
norleusina (rantai samping n-butil)




Kalkogen
Serina, homoserina, O-metilhomoserina, dan O-etilhomoserina memiliki rantai samping hidroksimetil, hidroksietil, O-metilhidroksimetil, dan O-metilhidroksietil; sedangkan sistein, homosistein, metionina, dan etionina memiliki ekuivalen tiol. Ekuivalen selenolnya adalah selenosistein, selenohomosistein, selenometionina, dan selenoetionina. Asam amino dengan kalkogen berikutnya juga ditemukan di alam: beberapa spesies fungi seperti Aspergillus fumigatus, Aspergillus terreus, dan Penicillium chrysogenum dalam ketiadaan belerang mampu memproduksi dan memasukkan telurosistein dan telurometionina ke dalam protein.[16]
Kode genetik yang diperluas
Peran
Dalam sel, terutama autotrof, beberapa asam amino non-proteinogenik ditemukan sebagai perantara metabolisme. Namun, terlepas dari fleksibilitas katalitik enzim pengikat PLP, banyak asam amino disintesis sebagai asam keto (seperti 4-metil-2-oksopentanoat menjadi leusin) dan diaminasi pada langkah terakhir, sehingga jumlah perantara asam amino non-proteinogenik tetap relatif rendah. [butuh rujukan]
Ornitina dan sitrulina terjadi dalam siklus urea, bagian dari katabolisme asam amino (lihat di bawah).[17]
Dimasukkan ke dalam protein secara pasca-translasi
Meskipun tidak dikodekan oleh kode genetik sebagai asam amino proteinogenik, beberapa asam amino non-standar tetap ditemukan dalam protein. Asam amino ini terbentuk melalui modifikasi pascatranslasi pada rantai samping asam amino standar yang ada dalam protein target. Modifikasi ini seringkali penting untuk fungsi atau regulasi protein; misalnya, pada γ-karboksilglutamat, karboksilasi glutamat memungkinkan pengikatan kation kalsium yang lebih baik,[18] dan pada hidroksiprolin, hidroksilasi prolin sangat penting untuk menjaga jaringan ikat.[19] Contoh lain adalah pembentukan hipusin dalam faktor inisiasi translasi EIF5A, melalui modifikasi residu lisin.[20] Modifikasi tersebut juga dapat menentukan lokalisasi protein, misalnya, penambahan gugus hidrofobik panjang dapat menyebabkan protein berikatan dengan membran fosfolipid.[21]
-
 Asam karboksilglutamat. Sedangkan asam glutamat memiliki satu gugus γ-karboksil, asam karboksilglutamat memiliki dua.
Asam karboksilglutamat. Sedangkan asam glutamat memiliki satu gugus γ-karboksil, asam karboksilglutamat memiliki dua. -
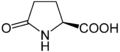
 Hidroksiprolina: Asam imino ini berbeda dari prolHidroksiprolin: Asam imino ini berbeda dari prolin karena adanya gugus hidroksil pada karbon 4.
Hidroksiprolina: Asam imino ini berbeda dari prolHidroksiprolin: Asam imino ini berbeda dari prolin karena adanya gugus hidroksil pada karbon 4. -
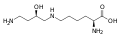 Hipusina: Asam amino ini diperoleh dengan menambahkan gugus 4-aminobutil (diperoleh dari spermidina) ke gugus ε-amino lisin.
Hipusina: Asam amino ini diperoleh dengan menambahkan gugus 4-aminobutil (diperoleh dari spermidina) ke gugus ε-amino lisin. -



Terdapat beberapa bukti awal bahwa asam aminomalonat mungkin ada, mungkin karena penggabungan abnormal, dalam protein.[22][23]
Analog toksik
Beberapa asam amino non-proteinogenik bersifat toksik karena kemampuannya meniru sifat-sifat tertentu dari asam amino proteinogenik, seperti tialisin. Beberapa asam amino non-proteinogenik bersifat neurotoksik dengan meniru asam amino yang digunakan sebagai neurotransmiter (yaitu, bukan untuk biosintesis protein), termasuk asam kuiskualat, kanavanina, karamboksin, dan asam azetidina-2-karboksilat.[24] Sefalosporin C memiliki kerangka asam α-aminoadipat (homoglutamat) yang diamidasi dengan gugus sefalosporin.[25]
-
 Tialisin
Tialisin -
 Asam kuiskualat
Asam kuiskualat -
 Kanavanina
Kanavanina -
 Asam azetidina-2-karboksilat
Asam azetidina-2-karboksilat -
 Sefalosporin C
Sefalosporin C -
 Penisilamina
Penisilamina



Lihat juga
Catatan
- ^ plus formilmetionin pada eukariota dengan organel prokariota seperti mitokondria
Referensi
- ^ Ambrogelly, A.; Palioura, S.; Söll, D. (2007). "Natural expansion of the genetic code". Nature Chemical Biology. 3 (1): 29–35. doi:10.1038/nchembio847. PMID 17173027.
- ^ Avan, Ilker; Hall, C. Dennis; Katritzky, Alan R. (22 April 2014). "Peptidomimetics via modifications of amino acids and peptide bonds". Chemical Society Reviews. 43 (10): 3575–3594. doi:10.1039/C3CS60384A. PMID 24626261.
- ^ Sarasa, Sabna B.; Mahendran, Ramasamy; Muthusamy, Gayathri; Thankappan, Bency; Selta, Daniel Raja Femil; Angayarkanni, Jayaraman (2020). "A Brief Review on the Non-protein Amino Acid, Gamma-amino Butyric Acid (GABA): Its Production and Role in Microbes". Current Microbiology. 77 (4): 534–544. doi:10.1007/s00284-019-01839-w. PMID 31844936.
- ^ Ostojic, Sergej M. (2021-08-01). "Creatine as a food supplement for the general population". Journal of Functional Foods. 83 104568. doi:10.1016/j.jff.2021.104568. ISSN 1756-4646.
- ^ Böck, A.; Forchhammer, K.; Heider, J.; Baron, C. (1991). "Selenoprotein synthesis: An expansion of the genetic code". Trends in Biochemical Sciences. 16 (12): 463–467. doi:10.1016/0968-0004(91)90180-4. PMID 1838215.
- ^ Théobald-Dietrich, A.; Giegé, R.; Rudinger-Thirion, J. L. (2005). "Evidence for the existence in mRNAs of a hairpin element responsible for ribosome dependent pyrrolysine insertion into proteins". Biochimie. 87 (9–10): 813–817. doi:10.1016/j.biochi.2005.03.006. PMID 16164991.
- ^ a b Lu, Y.; Freeland, S. (2006). "On the evolution of the standard amino-acid alphabet". Genome Biology. 7 (1): 102. doi:10.1186/gb-2006-7-1-102. PMC 1431706. PMID 16515719.
- ^ Voet, D.; Voet, J. G. (2004). Biochemistry (Edisi 3rd). John Wiley & Sons. ISBN 978-0-471-19350-0.
- ^ Chakauya, E.; Coxon, K. M.; Ottenhof, H. H.; Whitney, H. M.; Blundell, T. L.; Abell, C.; Smith, A. G. (2005). "Pantothenate biosynthesis in higher plants". Biochemical Society Transactions. 33 (4): 743–746. doi:10.1042/BST0330743. PMID 16042590.
- ^ a b c d e Weber, A. L.; Miller, S. L. (1981). "Reasons for the occurrence of the twenty coded protein amino acids". Journal of Molecular Evolution. 17 (5): 273–284. Bibcode:1981JMolE..17..273W. doi:10.1007/BF01795749. PMID 7277510. S2CID 27957755.
- ^ Koyack, M. J.; Cheng, R. P. (2006). "Design and Synthesis of β-Peptides With Biological Activity". Protein Design. Methods in Molecular Biology. Vol. 340. hlm. 95–109. doi:10.1385/1-59745-116-9:95. ISBN 978-1-59745-116-1. PMID 16957334.
- ^ Boniface, A.; Parquet, C.; Arthur, M.; Mengin-Lecreulx, D.; Blanot, D. (2009). "The Elucidation of the Structure of Thermotoga maritima Peptidoglycan Reveals Two Novel Types of Cross-link". Journal of Biological Chemistry. 284 (33): 21856–21862. doi:10.1074/jbc.M109.034363. PMC 2755910. PMID 19542229.
- ^ Arias, C. A.; Martín-Martínez, M.; Blundell, T. L.; Arthur, M.; Courvalin, P.; Reynolds, P. E. (1999). "Characterization and modelling of vanT: A novel, membrane-bound, serine racemase from vancomycin-resistant Enterococcus gallinarum BM4174". Molecular Microbiology. 31 (6): 1653–1664. doi:10.1046/j.1365-2958.1999.01294.x. PMID 10209740. S2CID 25796469.
- ^ Gao, X.; Chooi, Y. H.; Ames, B. D.; Wang, P.; Walsh, C. T.; Tang, Y. (2011). "Fungal Indole Alkaloid Biosynthesis: Genetic and Biochemical Investigation of the Tryptoquialanine Pathway in Penicillium aethiopicum". Journal of the American Chemical Society. 133 (8): 2729–2741. doi:10.1021/ja1101085. PMC 3045477. PMID 21299212.
- ^ Padmanabhan, S.; Baldwin, R. L. (1991). "Straight-chain non-polar amino acids are good helix-formers in water". Journal of Molecular Biology. 219 (2): 135–137. doi:10.1016/0022-2836(91)90553-I. PMID 2038048.
- ^ Ramadan, S. E.; Razak, A. A.; Ragab, A. M.; El-Meleigy, M. (1989). "Incorporation of tellurium into amino acids and proteins in a tellurium-tolerant fungi". Biological Trace Element Research. 20 (3): 225–232. doi:10.1007/BF02917437. PMID 2484755. S2CID 9439946.
- ^ Curis, E.; Nicolis, I.; Moinard, C.; Osowska, S.; Zerrouk, N.; Bénazeth, S.; Cynober, L. (2005). "Almost all about citrulline in mammals". Amino Acids. 29 (3): 177–205. doi:10.1007/s00726-005-0235-4. PMID 16082501. S2CID 23877884.
- ^ Vermeer, C. (1990). "Gamma-carboxyglutamate-containing proteins and the vitamin K-dependent carboxylase". The Biochemical Journal. 266 (3): 625–636. doi:10.1042/bj2660625. PMC 1131186. PMID 2183788.
- ^ Bhattacharjee, A; Bansal, M (2005). "Collagen structure: The Madras triple helix and the current scenario". IUBMB Life. 57 (3): 161–172. doi:10.1080/15216540500090710. PMID 16036578. S2CID 7211864.
- ^ Park, M. H. (2006). "The post-translational synthesis of a polyamine-derived amino acid, hypusine, in the eukaryotic translation initiation factor 5A (eIF5A)". Journal of Biochemistry. 139 (2): 161–169. doi:10.1093/jb/mvj034. PMC 2494880. PMID 16452303.
- ^ Blenis, J; Resh, M. D. (1993). "Subcellular localization specified by protein acylation and phosphorylation". Current Opinion in Cell Biology. 5 (6): 984–989. doi:10.1016/0955-0674(93)90081-z. PMID 8129952.
- ^ Copley, S. D.; Frank, E.; Kirsch, W. M.; Koch, T. H. (1992). "Detection and possible origins of aminomalonic acid in protein hydrolysates". Analytical Biochemistry. 201 (1): 152–157. doi:10.1016/0003-2697(92)90188-D. PMID 1621954.
- ^ Van Buskirk, J. J.; Kirsch, W. M.; Kleyer, D. L.; Barkley, R. M.; Koch, T. H. (1984). "Aminomalonic acid: Identification in Escherichia coli and atherosclerotic plaque". Proceedings of the National Academy of Sciences. 81 (3): 722–725. Bibcode:1984PNAS...81..722V. doi:10.1073/pnas.81.3.722. PMC 344907. PMID 6366787.
- ^ Dasuri, K.; Ebenezer, P. J.; Uranga, R. M.; Gavilán, E.; Zhang, L.; Fernandez-Kim, S. O. K.; Bruce-Keller, A. J.; Keller, J. N. (2011). "Amino acid analog toxicity in primary rat neuronal and astrocyte cultures: Implications for protein misfolding and TDP-43 regulation". Journal of Neuroscience Research. 89 (9): 1471–1477. doi:10.1002/jnr.22677. PMC 3175609. PMID 21608013.
- ^ Trown, P. W.; Smith, B.; Abraham, E. P. (1963). "Biosynthesis of cephalosporin C from amino acids". The Biochemical Journal. 86 (2): 284–291. doi:10.1042/bj0860284. PMC 1201751. PMID 13994319.
Konten ini disalin dari wikipedia, mohon digunakan dengan bijak.

